CHAPTER 3
The Botany of Citrus and Its Wild Relatives
BY WALTER T. SWINGLE
Revised by Phillip C. Reece
This account of
the citrus fruits and their numerous wild relatives has been written in the hope
of interesting both citrus growers and expert citrus investigators in this
large array of plants. Nearly every one of them has some striking
character—beauty of foliage; fragrance and beauty of the flowers; bright color
of the fruits; or, more practically, probable value as a rootstock for some of
the commonly cultivated species of Citrus or even possible use in
breeding new types of citrus fruits by hybridization.
The vast majority of citrus fruits and their
wild relatives are native to southeastern Asia, the East Indian Archipelago,
New Guinea, Melanesia, New Caledonia, and Australia; another group occurs in
tropical Africa. Many of the most interesting relatives of
Citrus have been collected only once or twice, and frequently flowers or
fruits are still imperfectly represented even in the largest botanical museums
of Europe and America. Consequently, it has been impossible to have
any consistent or uniform schedule for describing these plants in the present
chapter, but each genus has been given the best treatment that the material
available for study would permit. As a result, there is presented a
connected account of the whole orange subfamily and of the two tribes into which
it is divided, with some remarks on the geographical distribution and probable
evolutionary history of some of the more important genera of each of these
tribes. The thirty-three genera here discussed, however, have been
treated in as many essays varying somewhat in method, content, and taxonomic
technique with each genus.
Curiously enough,
the reader will find that the best-known material is often described with the
fewest words, whereas in an imperfectly known genus, in which the species are
still so inadequately known as to be difficult to separate without abundant
material, it has seemed necessary to describe each species in minute detail to
make sure that no possible differential character has been omitted.
It is hoped and believed that the reader will find something of interest in the
discussions given under each of the thirty-three genera.
Each tribe and every genus in the subfamily is
keyed out fully so that any of them can be easily identified if material is
available. Fourteen of the thirty-three genera contain only a
single species, so that the key to these genera in effect identifies these
species. The remaining nineteen genera, containing from two to many
species, are supplied with keys to determine all the species except with
respect to the genus Glycosmis, where only part of the species are keyed
out, although all are listed. It is much easier to get an idea of
the character of the species by studying the keys than by reading the detailed
descriptions, which are to be considered as material for reference.
These technical descriptions may, however, at any time become useful in the
event any little-known species becomes important either for ornament or for use
in practical citrus culture.
The commonly
cultivated citrus fruits belong to three genera, Citrus,
Fortunella, and Poncirus, all closely related and all belonging to
the subtribe Citrinae, of the tribe Citreae, of the orange subfamily
Aurantioideae, of the plant family Rutaceae. There are six other
subfamilies in the Rutaceae.
The family
Rutaceae, in turn, is obviously related to two other plant families: the
Simarubaceae, to which belongs the tree of heaven, Ailanthus altissima
(Mill.) Swing.; and the Meliaceae, of which the Chinaberry, Melia
azedarach L., is a well-known example. These plant families are
classed by taxonomic botanists along with some eighteen other families in the
natural order Geraniales, of which twelve families are included in the suborder
Geraniineae, which includes the Rutaceae.
Although the classification system of Engler is followed in the arrangement of
specimens in the larger herbaria in this country, it is not phylogenetic in the
modem sense. It follows a logical sequence of steps in the
arrangement of its larger taxa from the simple to the complex on the premise
that evolutionary lines progressed from apetaly to polypetaly and gamopetaly,
apocarpy to syncarpy, hypogyny to epigyny, and actinomorphy to
zygomorphy. Engler believed that flowers that appear simple have
always been simple. This view is now rejected by most botanists.
Hutchinson (1926) developed a presumed
phylogenetic system of classification based upon many of the principles on
evolutionary trends in the angiosperms adopted earlier by Bessey (1915) and
subscribed to by Fuller and Tippo (1949) and most students of floral
anatomy. The major point of divergence of the Hutchinson system
from earlier systems is the division of the dicotyledonous plants into two
subgroups: the Herbaceae composed of families that are predominantly
herbaceous, and the Lignosae whose members are predominantly
arborescent. Upon this premise the Rutaceae is placed in the
Rutales, an order Hutchinson considers derived from the
Celastrales. This system of classification places the Rutaceae in a
position quite remote from the Geraniales through which Engler and Diels
consider them derived.
Hutchinson's insistence
that seed plants had a monophyletic origin from hypothetical proangiosperms and
that families of predominantly herbaceous plants have evolved from the Ranales
and families of predominantly woody plants from the Magnoliales has prevented
the general acceptance of his system, but it has greatly stimulated phyletic
thinking in recent decades and may ultimately lead to the development of a
system based upon the works of Smith (1938) and Eames (1936) and an elaboration
of the skeletal system of classification of vascular plants set forth by Tippo
(1942).
The orange subfamily includes several
genera with rather numerous species that have a very primitive flower and
fruit structure, much like that of other subfamilies of the Rutaceae and even
analogous to that of several subfamilies in the plant families Simarubaceae and
Miliaceae, which stand very close to the Rutaceae in the natural system.
First of all, to show at a glance in clear
perspective the proper taxonomic placement of the orange subfamily in relation
to the six other subfamilies of the plant family Rutaceae to which it belongs,
an outline
of the Rutaceae is given showing all subfamilies and tribes and subtribes,
followed by a concise outline of the taxonomic arrangement of the orange subfamily
listing tribes, subtribes, subtribal groups, and genera, with page references
to each item.
It is worth noting that before
1943 no account of the orange subfamily describing all the genera and species
of the world had been published for more than a century. Prior to
publication of the original version of this chapter (Webber and Batchelor,
1943), the last complete account of the orange subfamily by a taxonomist was
published by Augustin P. de Candolle (1824, vol. 1, pp. 535-40) and contained
eleven genera and forty-three species, only a third as many genera and less
than a fourth as many species as are discussed here.
PRINCIPAL TAXONOMIC WORKS ON CITRUS AND RELATED
GENERA PUBLISHED SINCE 1860
In
December, 1860, Daniel Oliver read before the Linnean Society of London a paper
entitled "The Natural Order Aurantiaceae, with a Synopsis of the Indian
Species." It was published the following year (Jour. Linn. Soc.
Bot. 5 [2]: 1-44) and covered not only all the genera and species of this
order then known in India (nine genera and thirty-one species), but also
fourteen additional extra-Indian species belonging to six genera, including one
genus not found in India except in cultivation—a grand total of ten genera and
forty-five species. Oliver did not, however, attempt to treat the
genus Citrus. This was the first of the modem high-grade taxonomic
works on the orange subfamily.
In 1875, J. D.
Hooker recognized thirteen genera in the orange subfamily with forty-three
species (in Citrus he gave only four species) in the account of the
family Rutaceae in his Flora of British India (vol. 1).
In 1888, Dr. E. Bonavia brought forth a
voluminous treatise, The Cultivated Oranges and Lemons, etc., of India
and Ceylon, with an atlas published in 1890. This highly
imaginative work, by a genuine lover of plants, used for the most part only
native names for the varieties discussed, but it aroused interest all over the
world. Bonavia's theories of the morphology and evolution of citrus
fruits were highly original and ingenious, but departed widely from the
critical standards of professional morphologists and taxonomists.
In 1910, A. W. Lushington published "The Genus
Citrus" (Indian Forester 36:323-53) and gave names to many of the
Indian cultivated varieties studied by Bonavia. He also named some
of the citrus fruit trees figured and described by Rumphius in the seventeenth
century.
Henry N. Ridley, for many years
director of the gardens and forests (including the Singapore Botanic Gardens)
in the former British colony of Straits Settlements, published many articles
on rare Malayan plants, and a five-volume Flora of the Malay
Peninsula. Many new species of the orange subfamily were
described by Ridley in the first series of papers and coordinated with the
previously known ones in his Flora.
W.
G. Craib (1926, vol. 1, pp. 215-39), who wrote the Enumeration of the Flora
of Siam, discovered and described several very interesting new species of
Citrus relatives.
A. Guillaumin,
taxonomist of the Musée d'Histoire Naturelle at Paris, in 1911 gave a detailed
illustrated account of the Rutaceae in Lecomte's Flore genérale de
l'Indo-Chine. In the subfamily Aurantioideae he recognized
eleven genera and forty-four species, of which six were in
Citrus. He also published a valuable paper on the species of
Atalantia (in the broad sense) of continental Asia and on the
Citrus of New Caledonia (now placed in the genus Oxanthera).
I. H. Burkill, F.L.S., former director of the
Botanic Gardens at Singapore, has taken much interest in the species of
Citrus and related genera that are native in the Malayan
region. His "Enumeration of the Species of Paramignya,
Atalantia, and Citrus Found in Malay," published in 1931, and his
very valuable two-volume work, Dictionary of Economic Products of the Malay
Peninsula (1935), were used frequently in preparing this chapter.
E. D. Merrill, former chief of the Bureau of
Science at Manila, P.I., and later administrator of botanical collections and
director of the Arnold Arboretum, Harvard University, published much valuable
information regarding previously published species and described many new
species of Citrus relatives from the Philippines, Borneo, Indo-China, and
China.
Tyôzaburô Tanaka, who had specialized,
while a student in the Imperial University, on the study of the cultivated
varieties of Citrus, accompanied the senior author as guide,
interpreter, and assistant through the citrus-growing regions of Japan in 1915
(and again in 1918 and in 1926). He returned to Washington with
Swingle in 1915 and was employed as his assistant at different periods from
1915 to 1930 in the former Bureau of Plant Industry of the U.S. Department of
Agriculture.
In 1928-1930, Tanaka made a trip
around the world during which be photographed material of all the genera and
species of the orange subfamily to be found in the principal herbaria of Europe
and America. His studies of this material led to the publication of
a series of papers on the taxonomy of the subfamily Aurantioideae, entitled
"Revisio aurantiacearum." These papers constitute a very valuable
contribution to the taxonomy of this group. Tanaka's chief works
are concerned with satsuma varieties (Tanaka, 1932c) and citrus species
problems (Tanaka, 1954, 1959, and 1961).
In
1896, Dr. Adolph Engler published in the first edition of Die natürlichen
Pflanzenfamilien an account of the plant family Rutaceae. His
treatment of the orange subfamily included fourteen genera, and he estimated
the total number of species at about seventy-one, of which six were in the
genus Citrus. Thirty-five years later Dr. Engler, who for
more than fifty years had specialized on the Rutaceae, wrote, as his last
botanical contribution, a revised account of this family in the second edition
of Die natürlichen Pflanzenfamilien. He included twenty-nine
genera in the Aurantioideae and estimated the species at about 180, of which
eleven were in the genus Citrus.
Between 1912 and 1926, Walter T. Swingle (the senior author) published
seventeen taxonomic papers on tribes, subtribes, genera, and species of the
orange subfamily. Dr. Engler adopted nine of the genera Swingle
proposed and used his illustrations for several of them. It will be
evident to every reader of this chapter that Swingle has drawn freely on
Engler's great store of knowledge of the orange subfamily.
POSITION OF THE ORANGE SUBFAMILY,
AURANTIOIDEAE, IN THE PLANT FAMILY RUTACEAE
The family Rutaceae belongs to the division
Embryophyta Siphonogama, subdivision Angiospermae, class Dicotyledoneae,
subclass Archichlamydeae (Choripetalae and Apetalae), order Geraniales,
suborder 1, Geraniineae, along with eleven other plant families classed by
Engler and Diels (1936, p. xl) in the following order: Oxalidaceae,
Geraniaceae, Tropaeolaceae, Linaceae, Erythroxylaceae, Zygophyllaceae,
Cneoraceae, Rutaceae, Simarubaceae, Burseraceae, Meliaceae,
Akariaceae. The other suborders of the Geraniales are as follows:
2, Malpighiineae (3 families); 3, Polygalineae (2 families); 4, Dichapetalineae
(1 family); 5, Tricoccae (2 families); 6, Callitrichineae (1
family). The order Geraniales is preceded by the order Rosales
(including 17 families, among them Rosaceae and Leguminosae) and followed by
the order Sapindales (including 23 families, among them Anacardiaceae and
Sapindaceae).
The relationship of the orange
subfamily to the six other subfamilies of the
Rutaceae is shown by the general key given by Engler (1931, pp. 105-11),
which, translated somewhat freely from the German, reads as follows:
Subfamily I.
RUTOIDEAE. Carpels usually 4-5, seldom 1-3 or more, often united
only by the common pistil and free below, at maturity more or less separated,
opening inward by splitting the follicle (loculicidal), usually with a
dehiscent endocarp, very seldom with 4-1 fleshy drupes
(Pitaviinae). Leaves and bark of twigs with schizolysigenous oil
glands. (5 tribes, 17 subtribes, 86 genera.)
Tribe (i) Xanthoxyleae. Trees
or shrubs, mostly small, greenish or greenish-white (seldom large and clear
white) flowers which are always actinomorphous (radial) and often
unisexual. Carpels only seldom with more than 2 ovules, Embryo
[sic] mostly with flat cotyledons in endosperm (except Bosistoa
and Pagetia). (5 subtribes, 30 genera.)
Subtribe 1, Evodiinae (includes Xanthoxylon and Fagara) (20 g.),
tropics and subtropics of Old and New Worlds; subtr. 2, Lunasiinae ( 1 g.),
Monsoon region;1 subtr. 3, Decatropidinae (3
g.), Central America; subtr. 4, Choisyinae (5 g.), Central America, Pacific
Islands, and Australia; subtr. 5, Pitaviinae (1 g.), Chile;
Tribe (ii) Ruteae. Herbs or
perennial herbs, seldom shrubs, with medium-sized, always perfect flowers, which
sometimes (Dictamnus) are slightly zygomorphic. Carpels as a
rule with more than 2 ovules (only in Ruta in the subgenus
Haplophyllum with 2 ovules and occasionally indehiscent
fruitlets). Seeds with endosperm. (2 subtribes, 6
genera.) Subtr. 6, Rutinae (5 g.), subtropical and temperate
regions of Old and New Worlds (includes Cneoridium, a California shrub);
subtr. 7, Dictamninae (1 g.), temperate zone of Europe and Asia;
Tribe (iii) Boronieae.
Perennial herbs or shrubs. Carpels with only 1 or 2
ovules. Flowers always actinomorphic, mostly perfect.
Embryo usually straight, cylindrical, usually immersed in abundant fleshy
endosperm. (5 subtribes, 19 genera.) Subtr. 8,
Boroniinae (6 g.), Australia and New Caledonia; subtr. 9, Eriostemoninae (9
g.), Australia and New Caledonia; subtr. 10, Correinae (1 g.), Australia; subtr.
11, Nematolepidinae (2 g.), Western Australia; subtr. 12, Diplolaeninae (1 g.),
Western Australia;
Tribe (iv)
Diosmeae. Mostly perennial herbs and shrubs, seldom trees
(Calodendrum), always with simple leaves. Flowers almost
always actinomorphic, mostly perfect. Seeds without
endosperm. Embryo mostly straight with fleshy
cotyledons. (3 subtribes, 12 genera.) Subtr. 13,
Calodendrinae (1 g.) East Africa (Kenya) to Cape Province (South Africa);
subtr. 14, Diosminae (9 g.), Cape Province (South Africa); subtr. 15,
Empleurinae (2 g.), Cape Province (South Africa);
Tribe (v) Cusparieae. Shrubs
or tree. Flowers actinomorphic or the corolla and the androecium
zygomorphic. Seeds with little or no endosperm. Embryo
curved, the plumule lying between the cotyledons. (2 subtribes, 19
genera.) Subtr. 16, Pilocarpinae (3 g.), tropical America and
subtropical South America; subtr. 17, Cuspariinae (16 g.), tropical America,
mostly Brazil and northern South America.
Subfamily II. DICTYOLOMATOIDEAE. Leaves with
many-celled but not lysigenous oil glands. Flowers
actinomorphic. Stamens isomerous and alternate with the petals,
with bractlets at the base. Carpels united only at the base, with
several ovules. Fruits with dehiscent endocarp, 3-4
seeded. Small trees with doubly pinnate leaves. (1
tribe, 1 genus.)
Tribe (vi)
Dictyolomateae. (1 genus.) Brazil and eastern
Peru.
Subfamily III.
FLINDERSIOIDEAE. Trees or shrubs. Carpels 5-3,
united, each with 2-8 ovules in 2 rows. Fruit a loculicidal or
septicidal capsule, with persistent endocarp. Seed winged, without
endosperm. Leaves with lysigenous oil glands. (1 tribe,
2 genera.)
Tribe (vii)
Flindersieae. (2 genera.) Eastern Australia, New
Caledonia, East Indian Archipelago, Ceylon, and India.
Subfamily IV.
SPATHELIOIDEAE. Carpels 3, completely fused, each with 2 pendent
ovules. Fruit a winged drupe with a 3-loculed hard pit.
With oil-bearing secretory cells in the leaves, bark, and pith; lysigenous oil
glands in the leaf margins. (1 tribe, 1 genus.)
Tribe (viii) Spathelieae. (1
genus.) West Indies.
Subfamily
V. TODDALIOIDEAE. Carpels 5-2, incompletely or
completely united, or else only 1, each with 1 or 2 ovules. Fruit
formed out of 4-2 drupelets united only at the base, or which some occasionally
abort, or else a drupe with a thick or thin mesocarp and a thick or thin
endocarp, or a dry, winged, indehiscent fruit. Seeds with or
without endosperm. (1 tribe, 6 subtribes, 25 genera.)
Tribe (ix) Toddalieae. (6
subtribes, 25 genera.) Subtr. 18, Phellodendrinnae (2 g.),
temperate and subtropical eastern Asia and tropical Africa; subtr. 19,
Sohnreylinae (1 g.), Amazon Valley, Brazil; subtr. 20, Pteleinae (4 g.),
tropical and temperate America; subtr. 21, Oriciinae (2 g.), tropical Africa;
subtr. 22, Toddaliinae (13 g.), tropics, Old and New Worlds (include
Casimiroa, a Mexican and Central American fruit tree); subtr. 23,
Amyridinae (3 g.), northern South America, West Indies, Central America, Texas,
Florida, tropical Africa. [The genus Amyris, having about 30
species native to Florida, Texas, Mexico, Central America, West Indies, and
northern South America, is rather closely related to the tribe Clauseneae of the
next subfamily, Aurantioideae.]
Subfamily
VI. AURANTIOIDEAE. Fruit a berry [or hesperidium]
with a leathery rind or hard shell, in tribe Citreae often with pulp formed by
juicy emergenzen that arise on the carpellary walls. Seeds without
endosperm, sometimes with 2 or more nucellar [false] embryos.
Leaves and bark [of twigs and young branches] with schizolysigenous oil glands
[small or sometimes large trees, rarely shrubs.]; (2 tribes, 33
genera.) [This subfamily is given as classified by Swingle for
this chapter. Engler made a single tribe with 2 subtribes and with
a total of 29 genera.]
Tribe (x)
Clauseneae. (3 subtribes, 5 genera.) Subtr. 24,
Micromelinae (1 g.), Monsoon region and western Polynesia as far as Tonga,
Fiji, and Samoan Islands; subtr. 25, Clauseninae (3 g.), Monsoon region and
tropical Africa; subtr. 26, Merrilliinae (1 g.), Malay Peninsula and
Sumatra;
Tribe (xi)
Citreae. (3 subtribes, 28 genera.) Subtr. 27,
Triphasiinae (8 g.), Monsoon region; subtr. 28, Citrinae (13 g.), Monsoon
region and tropical Africa; subtr. 29, Balsamocitrinae (7 g.), Monsoon region
and tropical Africa.
Subfamily
VII. RHABDODENDROIDEAE. Flowers with a
bowl-shaped concave receptacle, with obliterated calyx, 5 petals, and very
numerous stamens. Ovary free, ovoid, 1-locular, with 1 basal
ovule. Pistil attached to the side of the ovary, with a long
lateral stigma. Fruits with thin exocarp and thin
endocarp. Leaves simple. (1 tribe, 1 genus.)
Tribe (xii) Rhabdodendreae. (1
genus.) Amazon Valley.
Total for
the Rutaceae: 7 subfamilies, 12 tribes (containing 29 subtribes), with about 150
genera and 1,600 species.
THE ORANGE SUBFAMILY,
AURANTIOIDEAE
The subfamily
Aurantioideae is defined briefly above in the outline of the Rutaceae, where all the subfamilies, tribes, and
subtribes of this plant family are given. All the species of the
Aurantioideae are trees or shrubs with persistent (evergreen) leaves except in
the three monotypic genera, Poncirus, Aegle, and Feronia,
and in three species of Clausena (C. pentaphylla, C.
dentata var. dulcis, and C. suffruticosa) and one of
Murraya (M. alternans). The flowers are usually white
and very often fragrant. Many of the genera bear subglobose fruits
with a green, yellow, or orange peel dotted with numerous oil glands that often
give an agreeable aroma when the fruit is handled. The fruits of
the genus Citrus are among the most beautiful, most fragrant, and most
delicious known to man. This subfamily contains Citrus and
thirty-two other genera related more or less to Citrus, classed in two
tribes and six subtribes which contain, as treated here, 203
species. The genus Citrus and a few others closely related to
it have fruits unlike any others known to botanists in being filled with
curious pulp-vesicles which contain in many species a delicious juicy
tissue. The subtribe Balsamocitrinae, which belongs to the tribe
Citreae, has fruits as large as oranges or grapefruits but with a hard woody
shell. These hard-shelled fruits do not contain juicy
pulp-vesicles, although some of them are pleasantly aromatic and much liked by
both natives and Europeans in India and Indo-China.
Many of the remote relatives of Citrus
belonging to the tribe Clauseneae have extremely small fruits very unlike those
of Citrus and usually semidry and entirely inedible.
Nevertheless some of these remote relatives have been found to be
graft-compatible with Citrus.
The
native habitat of the subfamily Aurantioideae is limited to the Old
World. Most of the genera are found in the Monsoon region from West
Pakistan to north-central China and thence south through the East Indian
Archipelago to New Guinea and Bismarck Archipelago, northeastern Australia, New
Caledonia, Melanesia, and the western Polynesian islands. Of the
thirty-three genera that constitute the Aurantioideae, no fewer than
twenty-nine are native to the Monsoon region and twenty-seven of them are found
only there. Five genera, belonging to two tribes and to three
subtribes, are native to tropical Africa and four genera are found only
there. Only one genus, Clausena, is native both to the
Monsoon region and to tropical Africa. Many of the species of
Citrus and of the genera closely related to Citrus are now found
in cultivation or are grown for ornament in all the tropical and subtropical
regions of the world.
Tribes, Subtribes, and Genera of the Subfamily Aurantioideae
The thirty-three genera of the subfamily
Aurantioideae are divided naturally and easily into two tribes: the
Clauseneae, with five genera, including the remote relatives of Citrus;
and the Citreae, with twenty-eight genera, including Citrus and all its
closer relatives. Each of these two tribes is divided into three
subtribes, making six in all. Keys are given to separate the two
tribes and also the subtribes [e.g., Clauseneae; Citreae].
As these tribes and
subtribes are frequently referred to in discussing relationships, English
phrases have been applied to them, in addition to their technical Latin names,
that will, it is hoped, suggest rather definitely the degree of relationship
of each tribe or subtribe to Citrus.
The natural order of the tribes and subtribes of the orange subfamily is from
the most simple to the most specialized, as shown in the list, except that the subtribe Balsamocitrinae, although
comprising species somewhat less highly specialized than those in the subtribe
Citrinae, has been placed at the end of the series. This has been
done to avoid intercalating the subtribe Balsamocitrinae between the subtribes
Triphasiinae and Citrinae, an arrangement which would have obscured the close
relationship clearly exhibited by some of the genera of Triphasiinae with some
of the more primitive genera of Citrinae. The list gives the number of groups under each subtribe, as well as
the number of genera. The genera having only one species
(monotypic) are marked with an asterisk; the number of species and varieties in
other genera is given after the genus name in parentheses.
TRIBE I. CLAUSENEAE: VERY
REMOTE AND REMOTE CITROID FRUIT TREES
The tribe Clauseneae includes the more primitive
genera of the orange subfamily. None of the species develop spines
in the axils of the leaves and the odd-pinnate leaves are at once distinguished
from those of the tribe Citreae by having the leaflets attached alternately to
the rachis, which does not break up into segments when the leaves fall;
rachises are not winged (except in Merrillia caloxylon, Murraya
alata, M. alternans, Clausena guillauminii, C.
wallichii,and C. luxurians).
Trifoliolate leaves are rarely found exclusively on any species but occur
sporadically merely by reduction of odd-pinnate leaves to trifoliolate leaves,
often on the same plant. Such trifoliolate leaves do not show
clearly the precise pairing of the lateral leaflets that is always shown in the
tribe Citreae. The leaves of plants of the tribe Clauseneae do not
show the winged petioles often found in members of the tribe Citreae (except
very narrow wings on the six species noted above).
The ovary has two to five locules with only one
or two ovules in each locule, except in Merrillia, which has five (rarely
six) locules and eight to ten ovules in each locule. The fruits are
usually small semidry or juicy berries, except in Merrillia, which has
ovoid fruits the size of a lemon, with a tough leathery peel. The
mature ovaries and young fruits of Micromelum have the locule walls
convolute. This genus has the petals valvate in aestivation,
differing thereby from all the other genera of the subfamily.
The bundle traces which enter the sepals and
petals in plants of the tribe Clauseneae were found by Albert H. Tillson (1938,
p. 9; Tillson and Bamford, 1938, p. 783) "to arise independently from the axis
and to display no trace of fusion with each other." In the tribe
Citreae only three of the twenty-four genera which he studied fail to show
such fusions.
Only a few genera
(five in all) have been established in this tribe, whereas the tribe Citreae has
twenty-eight. Seventy-nine species are listed here in the tribe
Clauseneae and 124 in the tribe Citreae, but the largest genus of the tribe
Clauseneae, namely, Glycosmis, for which thirty-five species have been
named, is still only imperfectly known.
The
tribe Clauseneae falls into three very natural subtribes, as is shown by the key.
Subtribe 1.
Micromelinae: Very Remote Citroid Fruit Trees
The subtribe Micromelinae contains only one
genus, Micromelum, with some nine or ten species, many of them very much
alike and rather hard to distinguish. All the species have
odd-pinnate leaves with alternate leaflets borne on a nonarticulated rachis,
except M. diversifolium, which has trifoliolate or unifoliolate
leaves. The inflorescences are corymbose-paniculate, often very
large and usually flat-topped, often surrounded by leaves much larger than even
the largest inflorescences.
The leaves of
Micromelum are very similar to those of Clausena and
Murraya, to which genera specimens of Micromelum are sometimes
assigned, even by experienced collectors and also by experienced
botanists. The flowers, ovaries, and seeds are, however, very
different. The petals of Micromelum are valvate, not
imbricate, and the cotyledons are flat and folded, not thick and plano-convex
as in all other genera of the subfamily Aurantioideae. The ovaries
show a curious twisting of the radial follicle walls, also unique in the orange
subfamily.
These remarkable diversities in
characters are of much taxonomic importance and are proof of the ancient origin
of Micromelum, a view also supported by the very wide distribution of
the genus in the Monsoon region, much farther to the eastward in a truly
indigenous state than any other genus of the orange subfamily.
I. Micromelum
Blume
I. Micromelum
Blume, Bijdr. Fl. Nederl. Indie 3:137. 1825; nomen conservandum,
Intern. Congr. Bot. 1930. Aulacia Lour. Fl. Cochinch. 1:273.
1790.
Type species.—Micromelum
pubescens Blume.
Distribution.—Northeastern India; Burma to Australia and New Caledonia;
Fiji, Tonga, and Samoan islands in western Polynesia.
Small spineless trees; leaves odd-pinnate (rarely
3-foliolate or 1-foliolate in M. diversifolium); leaflets thin, alternate
on wingless rachis; inflorescences usually large, terminal corymbose panicles
often flat-topped; flowers small, 5-merous (except the stamens, which are twice
as many as the petals); petals valvate in the bud; ovary with 2-6 (usually 3-5)
locules with 2 superimposed ovules in each locule; style rather slender,
narrowed at the base and articulated with the ovary, deciduous; radial walls of
the locules usually (always?) curved as if twisted during development by a
rotation of the outer wall of the ovary with reference to the axis; seeds with
thin, folded cotyledons; fruits subglobose or ovoid, dry berries with a
gland-dotted peel.
Oliver (1861, p. 19) gave
the following excellent sketch of the morphology of the genus Micromelum:
"…The essential characters of the genus [Micromelum] rest especially in
the broad, foliaceous, and remarkably contortuplicate cotyledons; perhaps,
also, in the singular torsion of the dissepiments of the ovary, which in M.
pubescens, is usually apparent immediately after the fall of the floral
whorls.…The fruit is but 1- (or 2-) seeded, and the twisted dissepiments with
the empty loculaments are soon closely pressed to one side by the young
seed. The style is minutely constricted and articulated at the
base ovary. In M. pubescens the ovary is very usually
5-locular; in Ceylon and Java specimens I have found an exceptional 6th cell;
in the Australian plant from Port Essington, etc., it varies, 4, 5 or
3. M. molle Turcz., I have found to be 4-locular, but
probably it also varies. The peculiar features which I have
remarked of the ovary and seed are associated with other common
characters.…These are especially the truly valvate or obliquely valvate
aestivation of the corolla, and the terminal cymose corymbs of numerous
flowers—as in Murraya seldom or never having the central axis of
inflorescence elongated as it is in the paniculate Clausenae…"
The genus Micromelum ranges from the
Samoan, Fiji, and Tonga islands to Australia and New Guinea, through the East
Indian Archipelago to the Philippines, Indo-China, southern China, Ceylon,
northeastern India, and West Pakistan. One species, M.
minutum, ranges farther eastward into Polynesia than any other species of
the orange subfamily and also occurs in New Caledonia, Australia, the
Philippines, and Indo-China. The other species have a more
restricted range, and one, M. ceylanicum, is restricted to the island of
Ceylon and another, M. diversifolium, is found only on Halmahera Island
and its near-by companion, Batjan Island, in the Moluccas.
THE SPECIES PROBLEM IN MICROMELUM
The number of species of Micromelum is not large—nine are here
recognized—but they can be described or keyed out only with difficulty because
apparently all the species are greatly variable, usually not merely in one, or
in a few, but in many characters. A cursory glance at the
literature of the genus shows that taxonomists are by no means agreed on the
number of species, or on the characters that define species, or on the areas
they occupy.
Fortunately, new characters are
being found that promise to make possible the satisfactory definition and the
easy recognition of the species when adequate material is
available. One of the most striking of these new characters is the
presence of large or small oil glands that are found, usually one above each
locule, at the tip of the ovary. Very large oil glands expand
greatly the ovary tip of M. falcatum; large oil glands occur in M.
compressum but do not noticeably expand the ovary tip; whereas only very
small oil glands are found similarly placed in M. pubescens.
These new characters, and others already used by
taxonomists, may be seen in figure 3-1, which shows microphotographs of longitudinal or cross
sections of ovaries of eight of the nine species of
Micromelum. A key to the species of Micromelum is presented.
1. Micromelum pubescens
Blume, Bijdr. Fl. Nederl. Indie 3:138. 1825. Illus. Engler, Die
Nat. Pflanzenfam. 3(4):186, fig. 107,A-K. 1896; fig. 3-1,D this work.
Type.—Java (Blume). Rijks Herb.,
Leiden.
Distribution.—Java, Sumatra,
Andaman Islands, southern Burma, Malaya, Palawan Island, P.I.
A brief description by Koorders (1912, vol. 2,
p. 424), based on material from Java, the type locality, reads, translated, as
follows: "Tree, up to 6 m high, or erect shrub. Leaflets 7-11,
alternate, soft-pubescent below, obliquely ovoid to broadly lanceolate, 3-17 X
2-7 cm. Fruit 6-8 mm long, orange-yellow." Ridley
(1922, vol. 1, p. 352) describes this species as follows: "Large shrub or
treelet. Branchlets and leaf-rachis puberulous but eventually
glabrescent. Leaves 6 to 18 in. long; leaflets 9 to 15, alternate,
lanceolate or ovate-acuminate, edges undulate, base slightly narrowed oblique;
nerves 9 to 12 pairs, 1.5 to 3.5 in. [3.8-8.8 cm] long, 0.5 to 2 in. [1.3-5 cm]
wide; petioles up to 3 in. [7.6 cm] long. Cymes 6 to 8 in. [15-20
cm] across, peduncled, many-flowered [pubescent]. Flowers 0.25 to
0.5 in. [6-13 mm] long, greenish-white. Calyx 5-toothed.
Ovary pubescent. Berry ovoid to oblong, glabrescent, orange, 0.3
to 0.4 in. [7.5-10 mm] long."
The chief
diagnostic characters of this species are given in the key. The
pistil (fig.
3-1,D) is of medium size with a rather long style in proportion to the
ovary. The oil glands at the tip of the ovary, one over each
locule, are unusually small and sometimes difficult to detect.
Tanaka (1931b, pp. 2, 3) contrasted this species with M. minutum,
as follows: M. pubescens has (1) much larger floral organs than M.
minutum; (2) stouter peduncles; (3) thicker leaflets; (4) ovary longer
stalked; (5) flower buds much broader and silvery pubescent, instead of
golden-yellow pubescent as in M. minutum. From M.
compressum of the Philippine Islands, Tanaka reported that it differs in
four characters (see next species below).
2.
Micromelum compressum (Blanco) Merr. Sp. Blancoanae 200. 1918. Bergera
compressa Blanco, Fl. Filip. 361. 1837; Micromelum tephrocarpum
Turcz. Bul. Soc. Nat. Mosc. 31:379. 1858. Illus. fig. 3-1,G.
Type.—Philippines, Luzon Island (Blanco). Original type
lost. Substitute type: (Merrill, Species Blancoanae, No.
884).
Distribution.—Philippines: Luzon
to Mindanao (and Palawan?) Islands.
The best
description yet published of this species is apparently Turczaninow’s original
diagnosis of his M. tephrocarpum, which, translated, reads as follows:
"Unarmed, very glabrous; leaves odd-pinnate, leaflets alternate, lanceolate,
obtusely acuminate [at the tip], attenuate, inequilateral at the based, entire
or slightly serrulate, pellucid-punctate; panicles terminal, much shorter than
the leaves; fruit ovoid-oblong, acute, becoming cinereous-blue, with fewer
than 3 seeds; calyx truncate, somewhat recurved, half as long as the petals;
alternate filaments shorter, anthers adnate, subglobose; ovary seated upon a
very short disk, style rather thick and terminated with a capitate stigma;
fruit (‘berry’) with the locule walls contorted and twisted
(‘contortuplicate’), 5-loculed, with only 3 seeds in the immature fruit [which
is] obovate-oblong, and flattened. From M. glabrescens
Benth. it seems to differ chiefly in having fruits with attenuate tips."
A flowering specimen collected by M. Ramos at
Bohol, Luzon, P.I. (Bur. Sci., No. 42570), in the herbarium of the Arnold
Arboretum, shows very small pistils only 2-3 mm long, with the ovary 1-1.2 mm
long, and the style about equaling the ovary in length, faintly furrowed
longitudinally, and abruptly contracted at the base where it joins the ovary,
about 0.4 mm diam., stigma hemispherical, 0.8 mm diam.
Tanaka, who has given much attention to
distinguishing the species of Micromelum and has examined carefully the
extant type specimens, has noted certain contrasts he observed between the
species. He stated (1931b, pp. 2, 3) that M.
pubescens, common in Java, Sumatra, and north to southern Burma, differs from M. compressum: (1) in having more slender
flower buds covered with very short silvery pubescence; (2) in its hairy ovary;
(3) in its glabrous pedicels; (4) in it's smaller, narrower leaflets with many
more lateral veins.
2a.
Micromelum compressum var. inodorum (Blume) Tan. Trans. Nat. Hist.
Soc. Formosa 22:418. 1932. Bergera inodora Blanco, Fl. Filip. 360.
1837; Micromelum molle Turcz. Bul. Soc. Nat. Mosc. 31:380.
1858.
Type.—Philippines (Cuming, No.
1056). Not located. Substitute type: (Merrill,
Species Blancoanae, No. 719). Herb. Bur. Sci.,
Manila.
Distribution.—Philippines;
doubtfully reported from Celebes by Tanaka (1932e, p. 419).
Turczaninow's original diagnosis of M.
molle (an undoubted synonym of Blanco's Bergera inodora) reads,
translated, as follows: "Unarmed; twigs, petioles, peduncles, panicles,
calyxes, and petals [covered with] short, soft pubescence; leaves odd-pinnate,
7-13-foliolate; leaflets alternate, ovate-lanceolate, acuminate [at the tip],
attenuate, inequilateral at the base, densely and softly pubescent on both
sides on the midrib and veins, sparingly pubescent between the veins; panicles
composed of both axillary and terminal [portions] shorter than the leaves;
calyx truncate, slightly 5-toothed, much shorter than the petals; fruits
acute, 1-3 seeded, blackish-blue, glabrous. From M. pubescens
Bl., of which a specimen was sent by Blume himself to the [Museum]
Schultesium, it differs in being more pubescent, in having larger and softer
[pubescent], acutely acuminate leaflets, and in having fruits acute, not very
obtuse."
Tanaka stated (1932e, p. 418):
"All botanical characters agree with the type species except the pubescence of
the leaf and the shoot." He also noted the short, thick flower buds
in contrast with the small, slender flower buds of M. pubescens and M.
minutum.
3. Micromelum
scandens Rechinger, Denkschr. Akad. Wiss. Math.-Naturw., Wien 89:564.
1914. Micromelum minutum var. intermedium Tan. Med. Rijks Herb.
Leiden 69:2. 1931. Illus. Tillson & Bamford, Amer. Jour. Bot.
25:782, figs. 1-11. 1938; fig. 3-1,F this work.
Type.—Bismarck Archipelago, New Britain Island, Gazelle Peninsula, Baining
Mountains (Rechinger, No. 3675). Herb. Naturhist. Mus.,
Vienna.
Distribution.—Bismarck
Archipelago and Melanesia.
The original
description reads, translated, as follows: "A climbing [or clambering] shrub;
leaves pinnate, leaflets large, 15-20 X 6-7 cm, with entire margins,
brownish-black when dried; fruits dry berries, disposed 30-40 in cymes, with
spirally twisted follicle walls, 8-12 mm long, glabrous, with many small pits,
black when ripe, with a short fragment of the style at the tip, and the flat
cupulate calyx, with an almost entire margin, persistent at the base."
The type specimen of M. scandens shows one
leaf 22 X 8 cm, larger than any yet measured of M. compressum, with 10
pairs of lateral veins; the leaflets of M. scandens are acuminate at the
tip and have 1/5-1/4 more lateral veins than those of M. compressum, and
they arise at a somewhat smaller angle with the midrib. The
persistent calyx is shallow, cupulate with indistinct lobes. The
pedicels of the fruits are 7-11 mm long, with 2 small bracts near the base.
A specimen in fruit collected on Bougainville
Island on May 25, 1930, by Kajewski (No. 1785) has a terminal fruit cluster
with very stout lower branches 2.5-3.5 mm in diameter. The notes
state that the fruits are "dark red when ripe, oblong coming to a blunt point,
length 1.4 cm, diameter 8 mm." Very young fruits are 3.5-4 X
1.2-1.5 mm, cylindrical, rounded at both ends and show a few scattered,
slender, white hairs. The nearly ripe fruits are glabrous, rough
with oil glands, and show at the tip a very short truncate style base 0.7-0.8
mm in diameter.
One of the specimens
collected by Kajewski in 1930 (No. 1654, from the Pupei Gold Fields,
Bougainville Island) was from a tree growing up to 18 m high. The
specimen has very large, lax, more or less leafy inflorescences with long
nodes below, much like those shown by the type specimen of the
species. The flower buds are oval or broadly and shortly
elliptical, 5-6 X 2-3 mm, decidedly larger than those of good material of
M. minutum from Tonga, the type locality, and from Fiji, New Caledonia,
and Australia. This specimen (Kajewski's No. 1654) had a flower
just ready to open; its pistil is 4.2 mm long when dry, ovary almost
glabrous. The calyx is large and expanded until it is shaped like a
shallow saucer, nearly pentagonal in outline.
Swingle considered M. minutum var. intermedium Tan. from New
Ireland, Bismarck Archipelago (to judge from a photograph of the types specimen
sent him by Tanaka), to be a fairly typical form of M. scandens, which
was based on a specimen collected by Rechinger in the adjoining island of New
Britain, the eastern-most point of which is distant only some 35 kilometers from
the west shore of New Ireland. Bougainville Island lies only some
200 kilometers southeast of the southern end of New Ireland. Tanaka
had not seen the excellent material of Micromelum from the Solomon
Islands in the Arnold Arboretum herbarium when he allocated just such a form to
M. minutum as a variety. Tanaka, who examined the type
specimen of M. scandens at Vienna, stated (1931b, p. 3) that it is
very different from M. minutum and most like M.
compressum. This species appears to be a good one, but needs
further study.
4. Micromelum
falcatum (Lour.) Tan. Bul. Mus. Hist. Nat. Paris, 2 sér. 2:157. 1930.
Aulacia falcate Lour. Fl. Cochinch. 1:273. 1790; Cookia
falcate DC. 1824; Micromelum octandrum Turcz. 1863. Illus.
fig. 3-1,C.
Type.—French Indo-China, Annam (?) (Loureiro). Herb. Brit.
Mus.
Distribution.—North Vietnam, South
Vietnam, Cambodia, Laos, Burma, Andaman Islands, Thailand, southern China.
Guillaumin (1911, p. 649) described this species
(under the name M. pubescens Bl.) as follows: "Shrubs or small tree,
twigs woolly-pubescent, then glabrescent. Leaves yellowish-green in
color, 15-31 cm long, odd-pinnate; leaflets 7-9, lanceolate, inequilateral at
base, long-acuminate-pointed, margins slightly crenulate, glabrous, except for
the midrib above and the larger veins below [which are pubescent], lateral
veins 6-8 on each side, prominent below, veinlets not visible; petiolules and
petioles cylindric, pubescent, 4-10 mm long. Inflorescences
pubescent, shorter than the leaves; pedicels woolly, 5 mm long, having two small
opposite hairy bracts toward the base; flowers 5 mm long, greenish-white; calyx
woolly with 5 very short teeth; petals 5, long-elliptic, glabrous or
glabrescent, 5 mm long; stamens 10, free, glabrous, the largest as long as the
petals; filaments subulate; disk short, glabrous, grooved; ovary ovoid,
velvety-hairy; style caducous, elongated, glabrous; stigma capitate; locules 5,
with 2 ovules in each; fruit ellipsoid, fleshy, with numerous oil glands,
glabrous; segments often 2-3, each with a single seed. Flowers
Dec.-Jan. Very common."
This
species differs from all others in the genus in having very narrow, often
falcate, long-acuminate leaflets, very unequal-sided at the base, usually 4 to
11 by 1.5 to 3 cm, including petiolule 3 to 5 mm long. It also is
the only Micromelum having the top of the ovary much broadened by large
oil glands, found one over each locule. This plant was the first
species of the genus to be described (as Aulaciafalcate in 1790), and
Loureiro's genus Aulacia antedates Blume's Micromelum by
thirty-five years. However, the genus Aulacia was not
understood by botanists in spite of the fact that a specimen of it, one of the
few remaining types of Loureiro's species, is preserved in the British Museum
at London. In 1919, Merrill (in a MS study on Loureiro's types)
concluded that Aulacia was a synonym of Micromelum, but the Fifth
International Botanical Congress, meeting at Cambridge in 1930, conserved the
name Micromelum and rejected the older generic name Aulacia
(Merrill, 1935, p. 221).
5. Micromelum ceylanicum Wight, Ill. Ind.
Bot. 1:109. 1840. Illus. fig. 3-1,A.
Type.—Ceylon (Wight, No. 1836). Herb.
Kew.
Distribution.—Found only in
Ceylon.
Trimen (1893, vol. 1, p. 218)
described this species (under the name M. pubescens) as follows: "A small
tree, shoots very finely and densely pubescent; leaves imparipinnate, 8-10 in.
[20-25 cm long], rachis pubescent; leaflets 9-15, alternate or subopposite,
shortly stalked, 1 1/2-3 in [3.8-7.5 cm long], very oblique base,
ovate-lanceolate attenuate, obtuse, irregularly finely crenate and wavy, with
very numerous, rather conspicuous glands, glabrous above, nearly so or slightly
pubescent beneath; flowers 1/4 - 1/2 in. [6-13 mm diam.], very shortly stalked,
arranged in terminal and axillary, spreading, pubescent, dichotomous,
carymbose cymes, bracts small, opposite at the bifurcations; calyx lobes broad,
triangular, pubescent; petals oblong-linear, spreading, pubescent; ovary
oblong, very pubescent; style rather longer, very thick, stigma capitate;
fruits 3/8 in. [9 mm] long, oblong-ovoid, pointed, rough with glands, glabrous,
yellow."
This species, as can be seen in figure
3-1,A has calyx lobes extending to the middle of the ovary in a nearly
full-grown flower bud. The other species of Micromelum would
not show the sepals in a section made at this level.
In Ceylon this species is said to be rather
common "in the low country, especially in the dry region." The name
of the species in Singhalese is walkarapincha. Murraya koenigii
is called karapincha in Singhalese and Clausena indica is
called mogen-karapincha. Both plants look so much like M.
ceylanicum as to be easily confounded with it, according to Trimen (1893, p,
219).
6. Micromelum
integerrimum (Buch.-Ham.) Roem. Syn. Hesper. 1:47. 1846. Bergera
integerrima Buch.-Ham. ex Coleb. Trans. Linn. Soc. London 15:367.
1827; B. integerrima Roxb. ex DC. nomen semi-nudem. 1824.
Illus. Beddome, Fl. Sylv. Anal. Gen. xliii-xliv, pl. 7 (flowers only).
1871; fig. 3-1,E this work
Type.—British India, Cult. Hort. Bot., Calcutta (Roxburgh).
Herb. Brit. Mus.
Distribution.—Northeastern India: West Bengal, Sikkim, Assam; Nepal; Burma;
East Pakistan; Andaman Islands.
Kurz (1877,
vol. 1, pp. 186, 187) described this species, under the name M.
pubescens, as follows: "An evergreen [shade-loving] tree, 25-30 ft.
[7.5-9.14 m] tall, with a clean trunk 10-12 ft. [3-3.66 m], measuring 2-3 ft.
[60-90 cm] in girth, all younger parts more or less densely puberulous; bark
about a line thick, whitish, somewhat roughish; cut pale-coloured; leaves
unequally pinnate, puberulous or glabrous, 1 1/2 ft. [46 cm] long, the rachis
usually more or less puberulous; leaflets in 3-7 pairs with an odd one,
alternating or nearly so, oblong-lanceolate to broadly lanceolate, oblique at
base, shortly petioluted, 5-7 in. [12-18 cm] long, acuminate, usually somewhat
waved, entire or nearly so; flowers middling-sized, whitish, on short pedicels,
forming terminal divaricate or crowded pale tawny appressed-pubescent
corymb-like panicles; calyx more or less obsoletely 5-teethed, pubescent;
petals about 2 1/2 lin. [5 mm] long, pubescent glabrescent; berries seated on
short, stalk-like torus, ovoid-oblong, also when young , glabrous,
gland-dotted, yellow, turning dull orange-red, terminated by the style
scar."
H. H. Haines, the former Conservator of
Forests of Bihar and Orissa (northeastern India) in his discussion of this
species (1921, p. 164), under the name M. pubescens stated: "A small
tree…leaves 8-18" [20-46 cm] long, with 5-11 very large leaflets and large
corymbs of white flowers 0.5" [13 mm] diam., which are succeeded by foetid,
ovoid, yellow or scarlet berries 0.5" [13 mm] long.…Leaflets ovate to
lanceolate or elliptical-oblong, attaining 8" by 3.5" [20 X 8.8 cm], lowest
sometimes only 1.5" [4 cm].…Corymbs pubescent or tomentose, often 1 ft. [30.5
cm] across…very pretty when in flower or fruit."
The flowers and especially the pistils of this
species are decidedly larger than those of all the other known species of
Micromelum. The fruits are attenuate at the base,
instead of broadly rounded as in M. hirsutum and several other
species. A specimen from the South Andaman Islands, secured by
King's collector in North Corbyu's Cove, now in the herbarium of the Arnold
Arboretum (Tanaka's Ident. No. A 64), shows (in the dry state) pistils 7 to 8
mm long; ovary 2 mm long; style 3 mm long with many fine longitudinal furrows;
stigma 1 mm high; the attenuate base of the ovary (disk?) 1.5 mm
high. Because of a long series of errors in citation, the botanical
name of this species has never yet been correctly accredited either as
Bergera integerrima or as Micromelum integerrimum.
Bergera integerrima was first published in 1827 by Colebrooke, who
accredited it to Buchanan-Hamilton. It was transferred to
Micromelum as a species distinct from M. pubescens by Roemer in
1846, a transfer incorrectly accredited to Wight and Arnott, who merely excluded
it from Bergera but did not recognize it as a valid species of
Micromelum.
7.
Micromelum hirsutum Oliv. Jour. Linn. Soc. Bot. 5(2):40. 1861.
Illus. Guillaumin, in Leconte, Fl. Gén. Indo-Chine 1:646, fig.
68(8-10). 1911; fig. 3-1,H this work.
Type.—Malay Peninsula, Penang (Wallich, No. 8516). Herb.
Kew.
Distribution.—Burma, Andaman
Islands, Thailand, North Vietnam, South Vietnam, Cambodia, Lao.
Oliver's original diagnosis of this species
reads in part, translated, as follows: "Leaflets 15-23, very asymmetrical at
the base; calyx small, with 5 triangular lobes; petals hirsute without; ovary
5-locular; stigma almost as long as the style; leaves 10-16 in. [25-41 cm
long]; leaflets trapezoid-lanceolate or ovate-lanceolate, often not a little
acuminate, somewhat obtuse at apex, margins slightly denticulate, sparsely
pilose or glabrescent below, pilose or hirsute particularly near the rib and
veins above. Inflorescences divaricate cymes, corymbose or
subpaniculate, hirsute. Pedicels very short. Petals
narrow or linear-oblong. Fruit (young) ovoid-oblong, hispid,
shortly and narrowly stipitate. Chiefly differing from M.
pubescens in the long leaves with very numerous leaflets, small calyx, and
petals strongly hairy externally."
Kurz (1877,
vol. 1, p. 187) added many details to Oliver’s technical diagnosis, as follows:
"A low, meagre, often simple-stemmed shrub, about 2-3 ft. [60-90 cm] high,
rarely higher, all parts more or less shortly tawny hirsute or puberulous
(rarely almost glabrous); leaves unpaired-pinnate, densely puberulous;
leaflets in 5-10 pairs with an odd one, lanceolate or oblong-lanceolate,
shortly but slenderly-petioluled, very oblique at base, acuminate, obsoletely
repand-serrate, usually 3-5 in. [7.6-12.7 cm] long; flowers small, whitish, on
short hirsute pedicels, forming more or less shortly tawny hirsute corymb-like
panicles at the ends of the branchlets and in the axils of the upper leaves;
calyx shortly tawny hirsute, deeply 5-lobed; petals hardly 2 lin. [4 mm] long;
ovary densely tawny hirsute; berries sessile or nearly so, oblong or obovoid,
especially while young more or less puberulous, gland-dotted, crowned by the
style scar."
"Hab.—Very frequent in the drier
upper-mixed forest, and the dry and low forests, all over Burma from Ava and
Pegu down to Tenasserim."
King (1894, p. 219)
added other details: "Leaves 6-12 in. [15-30 cm], rarely 15 in. [38 cm] long;
leaflets 9-25, 1.5-3.5 in. [4-9 cm] long, 0.8-1.5 in. [2-4 cm] wide; main
nerves 5-10 pairs. Cymes terminal, very tomentose, often lax, 6-8
in. [15-20 cm] in diameter but sometimes condensed and only 2 in. [5 cm] in
diameter."
This species differs chiefly from
the other species commonly occurring in Burma (M. integerrimum, but
called M. pubescens by King and Kurz) in being a small shrub, never a
tree, and in being more pubescent, especially the fruit, and in having smaller
flowers. Kurz noted that the fruits of M. hirsutum are
puberulous and almost sessile, whereas those of M integerrimum (his M.
pubescens) are short-stalked, glabrous berries. Guillaumin
(1911, pp. 649-50) noted that in Indo-China this species has "ellipsoid,
fleshy, reddish, glabrescent or pubescent fruits."
8. Micromelum minutum (Forst.)
Wt. & Arn. Prodr. Fl. Pen. Ind. Or. 1:94. 1834. Limonia minuta
Forst. f. Prodr. 33. 1786; Micromelum glabrescens Benth. Hook. Jour. Bot.
2:212. 1843; M. pubescens var. glabrescens (Benth.) Oliv.
1861; Glycosmis subvelutina F. Muel. 1858. Illus. Banks &
Solander, Bot. Capt. Cook's Voy. 1:14, pl. 36. 1900; partly reproduced in
fig.
3-2 this work; also fig. 3-1,B.
Type.—Friendly Islands (Forster). Herb. Brit.
Mus.
Distribution.—Samoa, Tonga, Fiji,
New Caledonia, northeastern Australia, Melanesia, Bismarck Archipelago, New
Guinea, and north to the Philippines, Sumbawa, and Borneo; also Annam
(fide Tanaka).
This species was
described, under the name M. glabrescens, in 1843 by Bentham from
material collected in the Friendly Islands (the type locality) by Barclay, as
follows: "Young leaves and inflorescences finely tomentose, leaflets 10-12,
obliquely ovate-acuminate, minutely crenulate, adult leaves glabrous; calyx
very short, 5-toothed; fruit oblong, very obtuse. This is evidently
very near M. pubescens Blume; does not quite agree with his [Blume’s]
very short description. The leaflets are quite smooth, except in a
very young state. The inflorescence is a dichotomous many-flowered
terminal cyme. The flowers appear very small, but are as yet
unexpanded in the specimen before me. The fruit is about 4 lin. [8
mm] long. The foliaceous cotyledons are very broad, deeply
emarginate and twisted, with rather a long straight radicle." An
examination of material from the type locality (Tonga Islands), from Samoa, and
from the Solomon Islands shows the following characters in the dry
state. The specimen from the Tonga Islands (coll. by H. E. Parks,
No. 16281, June-July, 1926, at Eua Island, now in U.S. Natl. Herb., No.
1528543) has very short pistils, only 2 mm long; the ovary is 1.3 mm long,
sparingly covered with short gray hairs; the style and stigma measure 0.8 mm,
the style is glabrous above, somewhat hairy below. The specimen
from the Solomon Islands (coll. by Kajewski, No. 2371, December 15, 1930, now
in Herb. Arnold Arbor.) shows pistils 3.5 mm long, with the ovary 1.5 mm long,
style and stigma 1.5 mm long. The ovary is fusiform, narrowed at
both ends, where it joins the disk below and the style above, and has scattered
yellowish hairs that soon fall off. The style is sparingly hairy,
not evidently furrowed longitudinally, sharply constricted at the base and
expanded into the flattened, discoid stigma, about 0.6 mm wide and 0.3 mm
tall. The specimens from Tutuila, Samoa (coll. by W. A. Setchell,
No. 257, now in Herb. Arnold Arbor.), show pistils 3 mm long; ovary 1.25 mm
long, covered with rather sparse, short, gray or yellowish hairs; style and
stigma 1.2 mm long; style with similar hairs at the base concealing the
constriction at the base, if present; stigma subglobose, 0.7 mm. diam.
This species has the smallest flowers and, in
particular, the shortest pistils of any species of the genus. It
ranges farther east in Polynesia than any other species belonging to the orange
subfamily. It alone reaches the Tonga and Samoan
Islands. It also ranges far to the north, from northeastern
Australia through New Guinea, Celebes, the Philippines, and reaches North
Borneo and Sumbawa, but not Java and Sumatra according to Tanaka (1931b,
p. 2), who stated, moreover, that Annam (Vietnam) is its westernmost limit.
As is to be expected from its very wide
distribution, this species varies considerably even in such important organs
as the pistil. Only two varieties have been published as yet, but
others could probably be made with equal justification.
8a. Micromelum minutum
var. tomentosum Tan. Trans. Nat. Hist. Soc. Formosa 22:419.
1932.
Type.—Timor (Forbes, No.
3753). Herb. Brit. Mus.
Distribution.—Timor; Philippines: Mindoro and Luzon islands.
Tanaka's original description of this variety,
translated, reads as follows: "Similar to Micromelum minutum in the
appearance and size of its flowers and fruits but all parts completely covered
with soft, golden yellow tomentum."
8b. Micromelum minutum var. curranii (Elm.)
Tan. Trans. Nat. Hist. Soc. Formosa 22:419. 1932. Micromelum
curranii Elmer, Leafl. Phil. Bot. 2:480. 1908; M. caudatum
Merr. Phil. Jour. Sci. 27:26. 1925.
Type.—Philippines, Luzon Island, Baguio (Elmer, No. 8530). Herb.
Bur. Sci., Manila.
Distribution.—Philippines: Bontoc, Benguet, and Union provinces of Luzon
Island, at 1,200-1,450 m altitude (fide Merrill, 1923, vol. 2, p.
335).
Elmer's original description of his
M. curranii (now reduced to a variety of M. minutum) reads as
follows: "Shrub, with rather numerous branchlets; wood hard, covered with
grayish-white mottled bark. Leaves crowded on the young twigs,
numerous, alternate, ascending, the peduncle [petiole] short yellowish
pubescent when young but soon becoming glabrous; leaflets alternatingly
scattered along the rachis, glabrous when mature, unequal in size, ovate to
broadly lanceolate, submembranous, entire or obscurely apiculate, apex
gradually acuminate, base obliquely rounded to subacute, drying brown, paler
beneath, the larger blades 7 cm long by 2 cm wide; petioles [petiolules] 3 to 5
cm long, when young short yellowish pubescent but becoming glabrous; nerves 3
to 5 pairs, ascending, more prominent beneath, reticulations not
visible. Corymbose panicle terminal, 6 cm wide across the top,
about as long; peduncles and pedicels strict, suberect, pubescent when in
flower, becoming glabrous when in fruit, the latter about 3 to 5 mm long; calyx
truncate or obscurely 5-apiculate, rim-like or saucer-shaped, pubescent when
young, persistent and glabrous in fruiting state; petals 5, ligulate, rather
thick, glabrous or sparsely pubescent on the outer side, 4 mm long, at least 1
mm wide, apex acute, deciduous; stamens in one series, about 9, inserted upon a
prominent disk at the base of the ovary, easily detached; filaments
subglabrous, fleshy and more or less flattened, pointed at the apex, subequal
in length, the longer ones 4 mm in length; anthers basifixed, broadly cordate
or subelliptic, nearly as wide; style thick, 1 mm long, bearing a terminal
subcapitate stigma, easily detached from the ovary; ovary glabrous, oblong in
outline or clavate, blunt at apex, base narrowed, immature fruits 1 cm long,
thicker above the middle, prominently punctate [of] lemon color, its [seeds
have the] cotyledons folded, dark green.…It is a form intermediate between
M. tephrocarpum Turcz. [M. compressum] and M. pubescens
Blm., but can be distinguished from either by its numerous, much smaller and
glabrous mature leaves."
9.
Micromelum diversifolium Miq. Ann. Mus. Bot. Lugd.-Bat. 1:211.
1863.
Type.—East Indian Archipelago,
Moluccas, Halmahera Island (Teijsmann). Rijks Herb.,
Leiden.
Distribution.—Known only from
the type locality.
Miquel's original diagnosis
reads, translated, as follows: "Twigs, petioles, and underside of the leaves,
as well as the inflorescence and the flowers, somewhat scurfy, slightly
tomentose-pubescent; leaves either with one pair of leaflets and an odd one,
thus trifoliolate, or unifoliolate leaflets, elliptic or oblong-elliptic,
obovate, acute or somewhat obtuse [at the tip], rounded or slightly emarginate
at the base, 7-8 spreading veins on both sides [of the mid-rib]; flowers in
short-terminal corymbs. Leaflets 2-4.5 in. [5-11.5 cm long]; calyx
with 5 short teeth; petals 5, valvate, pubescent without; stamens 10, the
alternate ones shorter, anthers ovate-cordate; style distinct, stigma convex;
berry ellipsoid, somewhat stipitate-constricted at the base with 5 locules,
with spirally twisted [radial] walls."
Tanaka
(1931b, p. 1), after having examined type specimens of this species in
Holland, both in the Rijks Herbarium at Leiden and in the herbarium at the
University of Utrecht, stated: "This extremely interesting species is not known
outside of the Moluccas. The most remarkable features are the
extreme reduction of the number of the leaflets, the exceedingly villose floral
organs and the completely tomentose berry."
The origin of a species of Micromelum found almost in the middle of the
area of distribution of the genus, differing radically from all the others in
the great reduction of the number of leaflets, is of much interest.
The unifoliolate form of the species (and of the variety mentioned below), has
reached the extreme reduction of the originally pinnate leaves to a single
leaflet that has occurred in species belonging to both the tribes of the orange
subfamily. Both tribes, however, still have species with
odd-pinnate leaves.
9a.
Micromelum diversifolium var. cuneata Miq. Ann. Mus. Bot.
Lugd.-Bat. 1:211. 1863.
Type.—The East Indies, Moluccas, Batjan Island (Teijsmann).
Herb. Univ. Utrecht.
Distribution.—Moluccas: Batjan, Obi, and Halmahera islands.
Miquel’s original diagnosis reads, translated,
as follows: "With larger glabrous leaflets [than those of the species], the
terminal one acute or cuneate at the base; the lateral leaflets somewhat acute
at the base with 8-10 ribs on both sides."
Tanaka (1931b, p. 1), after examining the type specimen in Holland, wrote
as follows: The "variety cuneata has a deep cupulate calyx and the
reduction of fuzziness is rather noticeable; it has occasional unifoliolate
leaves. Additional material from Obi and Halmaheira [Halmahera]
shows its fairly uniform nature."
Subtribe 2. Clauseninae:
Remote Citroid Fruit Trees
The subtribe
Clauseninae contains three genera, Glycosmis, Clausena,and
Murraya, having very simple, more or less primitive flower and fruit
structures. The flowers of Glycosmis, Clausena,and
Murraya are usually borne in dense, often large, panicled clusters at the
tips of the branches. None of the genera of this subtribe have
spines. The leaflets are alternate on the rachis, which is not
articulated and does not break into segments when the leaves fall.
The flowers are 3- to 5-merous except that the stamens (always free) are twice
as many as the petals. The ovaries have two to five locules and
each locule contains one to two ovules. The fruits are small
berries, either soft and juicy with few seeds immersed in mucilaginous pulp, or
semidry with a clearly distinguished, gland-dotted peel. The seeds
are glabrous.
The three genera,
Glycosmis, Clausena, and Murraya, included in the subtribe
range from India, Burma, and Indo-China to Borneo, the Philippines, New Guinea,
and Australia. Clausena also has a wide distribution in
tropical and subtropical Africa and in the bush forests, at high altitudes, in
the mountains of eastern and central Africa. Three species and two
varieties of Clausena are known from Africa. Plotted on a
globe, the distribution of the Remote Citroid Fruit Trees is very similar to
that of the Near-Citrus Fruit Trees.
Glycosmis and Clausena show very simple flower and fruit structures
much like those found in some genera of other subfamilies of the family
Rutaceae (such as Amyris in the subfamily Toddalioideae).
Close analogies to the flower and fruit structures shown by Glycosmis,
Clausena, and also Murraya can be found outside of the Rutaceae in
the Simarubaceae and Meliaceae, two plant families that stand very close to the
Rutaceae in the natural taxonomic system. The absence of oil glands
in the leaves and fruits is almost the only character that separates some of
the plants now classed in the Meliaceae from plants in the
Rutaceae. [A key to the genera of the subtribe Clauseninae is presented.]
The three genera of Remote Citroid Fruit Trees
are clearly though distantly related to Citrus. Citrus has
been grafted successfully on Clausena and Murraya and vice
versa. Such grafts are often short-lived, but sometimes live many
years. Grafts between Citrus and Glycosmis have not as
yet been successful, but so far only a single species of Glycosmis has
been tested, out of the large number known to exist. H. J. Toxopeus
(1936, p. 6) made hybrids between Citrus and Murraya, but they
were weak and remained stunted.
II. Glycosmis
Corrêa
II. Glycosmis
Corrêa, Ann. Mus. Hist. Nat. Paris 6:384. 1805. Sclerostylis
Blume, Bijdr. Fl. Nederl. Indie 1:133. 1825. Chionotria Jack,
Malay. Misc. 2(7):53. 1822; Dioxippe Roem. Syn. Hesper
1:33, 45. 1846; Myxospermum Roem. Syn. Hesper. 1:31, 40.
1846.
Type species.—Limonia
pentaphylla Retz. (Obs. Bot. 5:24 [1789]) = Glycosmis pentaphylla
(Retz.) Corr.
Distribution.—Southeastern Asia, East Indian Archipelago, Philippines, New
Guinea, northeastern Australia.
Unarmed small
trees or shrubs; new growth covered with dense, usually ferruginous pubescence;
leaves 1-foliolate, 3-foliolate, or odd-pinnate; leaflets with short petiolules,
alternate, long-lanceolate, more or less coriaceous; inflorescences compound,
often densely racemose; flowers small, 5-merous; sepals united to the middle,
with broad imbricate lobes; petals 5, white, long-elliptical or oval,
imbricate; stamens 10, filaments subulate, broadened below, anthers small,
often with oil glands on the back and tip; disk annular or cylindric; ovary
2-5-locular, with 1 ovule in each locule; style very short and thick,
persistent, stigma simple or disk shaped; fruit a juicy berry or dry; seeds
oval, thick, with a membranaceous testa; embryo with fleshy plano-convex
cotyledons and a very short plumule.
This
genus includes a large number of very closely related thornless shrubs or small
trees ranging from southeastern Asia and the East Indies to northern
Australia. It will probably require careful study of living plants
in the garden and in the laboratory to tell whether or not the very numerous
forms that have been referred to this genus constitute good species or are
merely varieties. Some species are constant, others
variable. Five species have uniformly unifoliolate leaves; two
have regularly trifoliolate leaves; and several have leaves with five leaflets;
two or three species occasionally have as many as 13 to 15
leaflets. Most of the species have very small flowers, but one
(G. macrantha Merr.), native to Borneo, is said by Ridley to have very
large flowers. Perhaps the most distinctive feature of Glycosmis
is the fact that the young twigs and leaves are densely covered with
closely-set, rusty-red pubescence, which does not seem to occur on any other
genus of Citrus relatives. The cotyledons of the seeds are
thick and fleshy like those of Murraya, Clausena,and most of the
other Citrus relatives, and differ widely from those of
Micromelum, which are thin and
folded.
Glycosmis citrifolia is
sometimes grown in greenhouses in this country, or even out of doors in the
southern states. The fruits are curious small berries having
translucent, slightly pinkish pulp, surrounding greenish-brown rounded seeds,
almost hemispherical when there are two seeds, but nearly spherical if there is
only one.
It is probable that, in addition to
G. citrifolia, other species are to be found in culture in European and
American greenhouses and gardens. Penzig (1887, pp. 194-209, Atl.
pls. 19-21) described and figured in detail the morphology and anatomy of two
forms of Glycosmis cultivated in Italy, one he determined as G.
pentaphylla Corrêa, the other with less certainty as G. lanceolata
Kurz. In this latter species, Penzig found the plumule of the
embryo, before germination, to have abundant, rather long, slender,
many-septate, brown hairs with punctate walls. Such hairy plumules
are exceedingly rare and were not known to Penzig anywhere else except in
certain species belonging to the Meliaceae, a plant family closely related to
the Rutaceae.
The species of Glycosmis
are still very inadequately described and need study more urgently than
those of any other genus of the orange subfamily.
TENTATIVE LIST OF SPECIES OF GLYCOSMIS
The taxonomy of Glycosmis is in such a
state of confusion that it is not yet possible to key out all the species or
even to tell with certainty how many should be recognized as valid.
Engler (1931, pp. 317-18) listed twenty-one
species as follows (numbers added and geographical names modernized): (1) G.
pentaphylla (Retz.) Corrêa (= G. cochinchinensis [Lour.] Pierre),
very widely distributed; leaves and inflorescences very variable; leaves one-
to two-paired or unifoliolate, with long-lanceolate, blunt or acuminate
leaflets; distributed in the whole Indian Malayan region through the Malay
Peninsula and Timor to northern Australia, and north to the Philippines (among
the many varieties is one with a single lanceolate leaflet, in the Khasi Hills,
Assam and Burma; another [G. simplicifolia Spreng.] occurs in Java); (2)
G. cyanocarpa (Blume) Sprengel, common in the Malayan region and also
found in the eastern Himalayan region by Hooker fils and Thompson and sent out
from Kew as G. arborea; (3) G. macrocarpa Wight, leaves with 5-1
leaflets; in southern India and Perak; (4) G. ovoidea Pierre, leaves with
5-1 leaflets; in Cambodia; (5) G. erythrocarpa Hayata; leaves with 3-1
leaflets; in Taiwan; (6) G. montana Pierre (= Tetracronia cymosa
Pierre, fide Tanaka), leaves with 13 leaflets; in South Vietnam
(Cochin China); (7) G. tomentella Ridley, leaves with 15 to 7 leaflets;
in Malay Peninsula, Selangor; (8) G. sapindoides Lindley, with a
pubescent ovary; in Andamans, Malay Peninsula, Java; (9) G. malayana
Ridley, leaves always 5-foliolate; abundant in Malay Peninsula from Singapore to
Perak; (10) G. monticola Ridley, leaves always 5-foliolate; rare; in
Malay Peninsula, Mt. Ophir, and Gunong Mering at 1,000 m altitude; (11) G.
elata Ridley, leaves always 5-foliolate; rare; in Malay Peninsula (Kelantan
and Kota Bharu); (12) G. mauritiana (Lam.) Tanaka (= Limonia
mauritiana Lam. [Ency., 3:51, 1789] = Limonia pentaphylla
Roxb. [Pl. Corom., p. 60, 1795] = G. triphylla Wt. [1833] =
G. nitida Wt. & Arn. [1834]), leaves with three or five leaflets,
staminal filaments flattened; along the Coromandel coast and in other parts of
India and Mauritius Island; (13) G. rupestris Ridley, leaves always
trifoliolate; in Malay Peninsula; (14) G. puberula Lindley, leaves always
trifoliolate; ovary pubescent; in Malay Peninsula.
The following species (nos. 15-18) all have
unifoliolate leaves: (15) G. dinhensis Pierre, South Vietnam (Cochin
China); (15a) G. bonii Guillaumin [= Murraya stenocarpa],
North Vietnam, Tonkin; (15b) G. pseudoracemosa [Guill.] Swingle [=
G. cyanocarpa (no. 2 above)], North Vietnam, Tonkin; (16) G.
crassifolia Ridley, Malay Peninsula; (17) G. lindleyana Swing. nom.
nov., G. macrophylla Lindley (non Miquel) ex Ridley [Fl. Malay
Penin., 1:349 (1922)], leaves unifoliolate, thin, elliptical,
acuminate, 8 by 3.5 inches; ovary conic, on a large disk; type, Wallich, No.
6377, Penang local; (18) G. parkinsonii Tanaka, Burma, Tenasserim.
Tanaka (1930a, pp. 47-49) reported fifteen
species in Indo-China and Thailand, among them: (19) G. pierrei Tanaka (=
Thoreldora cochinchinensis Pierre), South Vietnam (Cochin China); (20)
G. craibii Tanaka (= G. singuliflora Craib, non Kurz), Thailand;
(21) G. longipes (Craib) Tanaka, Thailand. Ridley (1930, p.
79) reported one new species from Borneo (21a, G. oliveri Stapf [=
G. macrantha Merr. (no. 29)]) with very large flowers. Engler
includes these species in his list.
In
addition to the foregoing list given by Engler (1931, pp. 317-18), Tanaka
(1928b, 1930a, 1930b, 1931b, 1932e, 1937) has
restored some old species and made a few new combinations and new species as
follows: (22) G. esquirolii (Lév.) Tanaka, China, Kweichow Province; (23)
G. parva Craib, Thailand; (24) G. winitii Craib, Thailand; (25)
G. bilocularis Thwaites, Ceylon and peninsular India; (26) G.
citrifolia (Willd.) Lindley, southern China, North Vietnam, South Vietnam,
Laos, Cambodia, Thailand; (27) G. chlorosperma Blume, Java, Malay
Peninsula; Borneo; Balabac Island, P.I.; (28) G. clemensii, Tanaka, North
Borneo, Mt. Kinabalu; (29) G. macrantha Merrill, Borneo, Tawau; (30)
G. angularis Elmer, Leyte Island, P.I.; (31) G. greenii Elmer,
Philippines, Borneo, Moluccas, New Guinea, Australia; (32) G. platyphylla
Merrill, Leyte and Samar islands, P.I.; (33) G. singuliflora Kurz, India,
upper Assam; (34) G. macrophylla (Bl.) Miquel, Moluccas, Kei Islands;
(35) G. boreana Narayanaswami, India, Assam.
Tanaka also transferred G. bonii (no.
15a above) to the genus Murraya (as a synonym of M.
stenocarpa) and considered G. pseudoracemosa (no. 15b) to be a
synonym of G. cyanocarpa (no. 2) and G. oliveri (no. 21a) a
synonym of G. macrantha (no. 29). This makes the tentative
list total thirty-five supposedly valid species. There are also
many varieties both old and new not listed here.
To show how many apparently good species can be
made out of a single species of the older authors, Kurz (1876, pp.-33-40) made
ten species that he thought good out of what had been called G.
pentaphylla (Retz.) Corrêa by Oliver (1861) and Hooker (1875).
Kurz described all ten and illustrated seven. His key gives an
excellent idea of the characters shown by the species of Glycosmis; it
is reproduced with slight emendations:
Of the foregoing ten species segregated by Kurz
from G. pentaphylla (fig. 3-3), as understood by Oliver (1861) and Hooker (1875), four
are listed by Kurz as having named varieties: (a) G. cyanocarpa
and (d) G. trifoliata, each with four varieties; (e)
G. triphylla and (h) G. arborea, each with two varieties in
addition to the typical form. Seven of these segregates,
a, b, c, e, f, g, h, of the key
presented are figured in two plates.
Of these
ten species, Kurz considered f, i, and k as distinct species
"beyond any doubt," but was undecided whether d, e, and g
should not be thrown "all into one species." In the preceding
numbered list, six of Kurz's species are included: a = no. 2; c =
no. 1; e = synonym of no. 12; f = no. 33; g = no. 14;
k = no. 17. One species reduced to varietal rank by Kurz
(var. beta of species a) was considered a good species by Engler (no. 8
above).
It is very necessary that all the
species and varieties of Glycosmis be subjected to a critical revision
and then be fully described and provided with a workable key. The
descriptions of the older species do not suffice to separate these species from
the many new ones now recognized, some of which are also inadequately
described.
One or more illustrations of the
following eleven species of Glycosmis are cited in Stapf (1929-1931, vol.
3, p. 298, and vol. 6, p. 554): G. arborea var. insularis Kurz;
*G. cochinchinensis Pierre (no. 1); G. cyanocarpa Spreng. (no. 2);
G. cymosa Kurz; G. lanceolata Spreng.; *G. madagascariensis
Corrêa; G. montana Pierre (no. 6); G. pentaphylla Corrêa (no.
1), "(sub nomen Limonia pentaphylla)"; G. puberula Lindl. (no.
14); G. singuliflora Kurz (no. 33); *G. triphylla Wight (no.
12). (The species names preceded by an asterisk have colored
illustrations and those followed by a number in parentheses are thereby assigned
to the species in the numbered list above.)
Glycosmis citrifolia (Willd.) Lindl.
Trans. Hort. Soc. Lond. 6:72. 1826. Limonia citrifolia Willd.
Enum. 448. 1809; L. parviflora Sims, Bot. Mag. pl. 2416. 1823;
Glycosmis pentaphylla, var. beta, subvar. 2 (chinensis) Oliv.
Jour. Linn Soc. Bot. 5(2):37. 1861. Illus. Sims, loc. cit.
pl. 2416; Tillson & Bamford, Amer. Jour. Bot. 25:782, figs. 12,
13, 1938; Everett, Addisonia 21:pl. 687.
1940.
Type.—(Willdenow, No. 8116),
Herb. Bot. Mus., Berlin-Dahlem.
Distribution.—Southern China, North Vietnam, South Vietnam, Cambodia, Laos,
Thailand; widely cultivated in the Old and New
Worlds.
Common name.—Chinese
glycosmis.
A small tree or shrub with slender,
glabrous, greenish twigs with nodes 2-4 cm apart; leaves 1-, 2-, or
3-foliolate, rarely 4- or 5-foliolate; leaflets (or the single blade)
elliptical or oblong-elliptical, 6-17 X 2-5 cm, usually acute at the tip and
narrowly cuneate at the base, rarely somewhat rounded at base, more rarely at
tip, more or less covered with rusty-brown, short pubescence when very young
but soon glabrescent, lateral veins very numerous, 12-15, visible on both
surfaces, variable in size, only the stronger ones running nearly straight
about 3/4 of the way to the margin, at an angle of 50°-60° with the midrib;
petioles 1.5-2.5 cm long, articulated with the blade in simple leaves;
petiolules of lateral leaflets 4-8 mm long, with a channel on the upper side
made by the decurrent leaf margins, articulated only at the base with the
rachis; the terminal leaflet of pinnate leaves often articulated with a free
portion of the rachis 1-2 cm long; inflorescences small, axillary, usually 2-3,
sometimes 4-5 cm long, rarely terminal, sparingly branched, pedicels very
short, all parts of the inflorescence covered, when young, by a dense
rusty-brown pubescence; flowers small, white, 4-5-merous; sepal lobes small,
subtriangular, with more or less ciliate margins; corolla campanulate, petals
white, small, 3.5-4.5 mm long, bluntly cuneate at the tips, soon falling;
stamens 10, filaments filiform, slightly dilated upward; ovary short, ovoid,
1.5-2 X 1.3-1.6 mm, rough or tubercular, merging into a short, thick,
persistent style, 0.4-0.5 mm long, tapering upward and ending in a depressed
cushion-shaped, persistent stigma, 0.5 mm wide and 0.2-0.3 mm high; fruits
subglobose, translucent white or pinkish, 11-13.5 mm diam.; seeds 1-3, oblong,
cotyledons green.
This species, the Chinese
glycosmis, although widely cultivated in all tropical countries and in
greenhouses in the temperate zones, is still inadequately studied.
The description above was based on the type specimen (leaves only) preserved at
the Berlin-Dahlem herbarium, and on material from southern China determined as
this species by Tanaka. The Chinese glycosmis is often labeled
G. pentaphylla (Retz.) Corrêa in gardens or herbaria, as many authors
have considered it as being merely a form of that species. It has
also been called G. cochinchinensis Pierre by some botanists.
These three species, and all the others for that matter, should be introduced
into the United States and grown side by side so that they can be properly
distinguished, adequately described, and tested as possible rootstocks for
Citrus and as ornamental plants for greenhouse culture or for planting in
subtropical gardens and parks. Glycosmis citrifolia has
become naturalized in the hammocks of Key West, according to Small (1933, p.
759) and Everett (1940, p. 29).
III. Clausena Burm.
f.
III. Clausena Burm. f.
Fl. Ind. 87, 243. 1768. Cookia Sonner, 1782; Fagarastrum G. Don.
1832; Gallesioa Roem. 1846; Myaris Presl. 1849; Quinaria
Lour. 1790; Piptostylis Dalz. 1851; Polcyema Voigt.
1845.
Type species.—Clausena
excavata Burm. f.
Distribution.—Southwestern Asia, East Indian Archipelago, Philippines, New
Guinea, Australia, northeastern tropical Africa from Abyssinia south to Natal
and Pondoland in Cape Province of the Republic of South Africa, central tropical
Africa, and western Africa from Angola to Sierra Leone.
Trees or shrubs without spines; leaves
odd-pinnate, leaflets alternate, usually 3-7 sometimes 19-32; rachis usually
not winged; inflorescences terminal or axillary, paniculate or laxly racemose,
flowers often in cymose clusters, flower buds small, subglobose or short-oval
or oblong, never long-cylindric; flowers small, calyx 4-5 lobed; sepals fused
into a cup below; petals free, 4 or 5 imbricate in the bud, usually elliptical;
stamens 10, in 2 whorls, the outer row opposite, the sepals usually longer,
filaments more or less dilated or flattened below, often geniculate where the
filiform apical portion joins the flattened and expanded basal portion, anthers
ovate or elliptical, rarely short and subglobose; gynophore well developed,
usually a perfectly glabrous hourglass-shaped column resting on an annular
nectary below and supporting (sometimes clasping) the base of the ovary above;
ovary with 2-5 locules, often pubescent or glandular, ovules 2 in each locule
(rarely 1); style deciduous, often shorter than the ovary or equaling it in
length (rarely longer), thick, sometimes merging gradually into the ovary,
often sharply delimited and narrowed where it joins the ovary, stigma
inconspicuous, sometimes subcapitate after the style shrivels; fruits small,
subglobose or ovate, with 2-5 segments, sometimes only 1 seed maturing; all
flower parts usually showing many (not all) cells strongly impregnated with
tannin residues.
The most distinctive
morphological character of the genus Clausena is the gynophore, which in
the typical species is a large, well-developed, hourglass-shaped structure
supporting the ovary. It is perfectly glabrous and rests upon and
merges into a short annular nectary below and sometimes expands above into a
thin-lipped cup-like structure that encloses the base of the ovary.
However, the gynophore varies greatly in the different groups of species and in
some is so modified by expansions caused by development of oil glands that it
is difficult to recognize. Nevertheless, it is present in all
species of Clausena and separates them from the species of other related
genera.
RELATIONSHIPS OF THE SPECIES OF CLAUSENA WITH ONE ANOTHER
The numerous species of this genus, still only
imperfectly studied with respect to the minute flower characters, cannot be
arranged now in natural sections or subgenera. It is clear to any
taxonomist who looks over carefully a large collection of the species of
Clausena that they can be classified into obviously related groups, some
small, some large. Unfortunately, these groups are not clearly
distinguished from one another but, on the contrary, seem rather to merge into
allied groups without any sharp line of demarcation.
The study of the flower characters and in
particular of the gynoecium by means of serial microtome sections has shown
that important characters are found in the pistil that may help to define the
natural groups of species.
The type species of
the genus, C. excavata, has a striking hourglass-shaped gynophore which
is completely glabrous. It arises from the annular nectary below
and, after contracting, again expands and may even be slightly flaring where
it joins the ovary, which is strongly hirsute. The ovary, which is
5-locular, merges into the style without any clear line of demarcation, and the
style is not narrowed where it joins the tip of the ovary.
On the other hand, in C. indica the style
is abruptly contracted and countersunk into a conical depression at the tip of
the ovary, which has two to five locules. This species was made the
type of the genus Piptostylis and belongs to a large group which includes
many other species.
The species C.
pentaphylla at present prevents a clear separation of the two groups
outlined above. The pistil shows no sharp boundaries of gynophore,
ovary, and style; the gynophore is as broad as the ovary—not
hourglass-shaped—and is more or less lobed, often having short processes each
ending in an oil gland. The style may be slightly contracted where
it joins the top of the ovary but is frequently more or less swollen with oil
glands just at this junction.
Clausena lansium stands apart from all the other species of
the genus in having a star-shaped bud (due to its five strongly carinate
petals) and a 5-angled ovary, corresponding in position to the five petal keels
in a cross section of the bud. The pistil is unusually large; the
ovary is very strongly hirsute and merges into the style, which is slightly
contracted at the base. It is hourglass-shaped as in C.
excavata. Furthermore, the twigs show a central gum canal,
doubtless formed by a lysigenous breakdown of the tissues. The
locules of the ovary contain scattered but well-developed hairs which arise on
the dorsal and outer portion of the lateral walls and usually grow more or less
toward the axis of the ovary. One much longer hair arises at the
base of each locule and grows vertically upward, nearly to the top of the
locular cavity (Penzig, 1887, p. 179, pl. 16, figs. 11, 12, 16).
The fruits are apparently the largest in the genus and the locules of the ripe
fruits are almost filled with a pulpy tissue which Penzig found to arise from
the hypertrophy and multiplication of layers of cells nearest to the
endocarp. This species was the type of the genus Cookia, but
no other species having similar characters are yet
known.
Clausena lenis also stands alone in the genus, having
leathery petals and a most peculiar gynophore, no longer hourglass-shaped as in
almost all the other species. Just above the thin annular nectary
there is a narrow constriction followed by a cylindrical gynophore evidently
swollen by a ring of 12 to 15 oil glands at its very base; it merges above
into the ovary. The style is slender, two to three times as long as
the ovary, by far the longest of any species in the genus, and contracted
below, where it joins the ovary. The leaflets are subdentate, and
the largest ones are lateral leaflets attached about two-thirds of the way up
the rachis.
Clausena guillauminii has
leaves showing dimorphic oil glands, the larger ones being 1/3 to 1/2 mm in
diameter, and deep red by transmitted light; the flower
parts show a single large oil gland at the tip of each sepal, each petal, each
anther, and each locule of the ovary. C. wallichii has almost
the same distribution of the oil glands in the flower parts. C.
papuana shows great variation in the size of the oil glands in the leaflets
and has the oil glands similarly distributed in the flower parts.
These species may be closely related; certainly C. guillauminii, C.
wallichii and C. luxurians are. The last-named species
have the rachis slightly winged, unlike all other species of
Clausena.
THE SPECIES PROBLEM IN CLAUSENA
Of
the twenty-three species of Clausena recognized here, ten were published
from 1900 to 1930 and one as late as 1940. Of the others, one was
published in 1892 and eleven from 1768 to 1875. It now appears that
many of the older species, although they were described adequately enough to
distinguish them from the other species known at the time, were nevertheless
not described fully enough to separate them clearly from the many species
recently discovered. Then, too, the range of the older species has
been extended steadily as new botanical collections have been made.
Very widely distributed species like C. excavata and C. anisata
show, as might be expected, very many variations in their characters in
specimens collected from widely different localities.
For these reasons a clear-cut definition of the
species of Clausena will require a great amount of work which can be done
only by using all the material in the leading herbaria of the world.
Tanaka, who assiduously studied Clausena
(and also the other genera of Aurantioideae) from 1927 to 1937 in the
herbaria of Europe, Asia, and America, has done much to clarify the species of
this genus. He has published notes on them in all his valuable
papers included in the series "Reviso aurantiacearum."
Swingle was permitted to borrow nearly a thousand
sheets of material of Clausena from many of the leading herbaria of the
Old World and the New, including all the Clausena material in the
herbariums of the Royal Botanic Gardens at Kew and the Royal Botanic Garden at
Calcutta. Thanks to these loans he was able to make a close study
of many of the actual type specimens of the species of Clausena.
In the study of this rich material he used a modification of Juel's method
of softening herbarium material, after which it was fixed, dehydrated,
imbedded, cut into serial microtome sections, double-stained, and finally
mounted permanently on glass slides (Swingle, 1939b, p. 270).
In this work, Swingle had the skillful assistance
of Dr. Albert H. Tillson. Figure 3-4, which shows longitudinal microtome sections of nine
species of Clausena, makes clear how helpful this method is in studying
herbarium material, especially the pistil just after the petals and stamens
have fallen. It has not, however, been possible to work out all
the species satisfactorily. Some species, like C. anisata,
vary so greatly that months of work should be needed to study its numerous
forms adequately, and other species are not well enough represented in herbaria
for any complete study.
GEOGRAPHICAL DISTRIBUTION AND USES
The geographical distribution of Clausena is of interest since it has the
widest range of any genus included in the orange subfamily, species being found
all the way from northwestern India to China and Taiwan, south through the East
Indian Archipelago to Timor, northern Australia, and New Guinea.
Moreover, there is a group of three species that covers a wide range in Africa,
from Ethiopia (Abyssinia) to Cape Province and through western Africa from
Angola north to Sierra Leone.
There are wide
differences in the character of the growth and the height of the species; they
range all the way from shrubs 20 to 40 cm high in Indo-China to trees reaching
a height of 20 m (66 feet) in Africa.
Several
species of Clausena produce edible fruits, but up to now only one of
them, the Chinese wampee, C. lansium, is cultivated (in southeastern
China) on a commercial scale, with many varieties, the fruits varying greatly
in size, flavor, and time of ripening. One other species, the
Indian wampee, C. dentata var. dulcis, is gathered on a large
scale from wild trees in the state of Madras in southern India, where it is a
highly appreciated fruit, said by the British experts who have studied it to be
of excellent flavor. At least two other forms are said to produce
edible fruits, C. indica of India and Ceylon and C. dentata var.
henryi of southwestern China.
The
Chinese wampee, C. lansium, can be used as a rootstock for Citrus, provided a few twigs of the wampee
are left growing below the graft. Other species of Clausena
should be tested as stocks for Citrus and also for the edible-fruited
species of Clausena.
Clausena
anisum-olens, a species common in the Philippines from Luzon Island to
Mindanao Island, grows in the forests from sea level to 1,500 m
altitude. It is a small tree 3 to 6 m tall and has leaves with a
strong odor of anis oil. West and Brown (1920, pp. 211-12) stated
that the suggestion had been made that the leaves could be used locally in
preparing anisado, a favorite alcoholic beverage among the
Filipinos. The essential oil of the leaves was found by Brooks
(1911, p. 344) to consist very largely (from 90 to 95 per cent) of methyl
chavicol, which is easily converted into anethol, the true anis
oil. The leaves yield about 1.2 per cent of oil after five hours’
distillation over steam. This essential oil is not known to occur
in such a large proportion in any other plant of the family Rutaceae.
It is highly probable that other species of this
large and very widely distributed genus, when studied carefully, will be found
to be of value for their fruits, for use as rootstocks, for their essential
oils, or for planting as ornamentals because of their abundant white flowers
and handsome foliage.
The following four keys
covering the species native to the four principal regions where Clausena
is indigenous will help in giving an idea of the differences between the
species.
1. Clausena excavata Burm.
f. Fl. Ind. 87. 1768. Murraya burmanni Spreng. 1825; Amyris sumatrana
Roxb. 1832; A. punctata Roxb. 1832; Gallesioa graveolens Roem.
1846; Clausena lunulata Hay. 1911; C. tetramera Hay. 1916; C.
moningerae Merr. 1923. Illus. Burmann f. loc. cit. pl. 29,
fig. 2; Guillaumin, in Lecomte, Fl. Gén. Indo-Chine 1:664, fig. 70(8-10).
1911; Tillson & Bamford, Amer. Jour. Bot. 25:782, fig. 15. 1938; fig.
3-4,A-B, and fig. 3-5 this work.
Type.—Java (Burmann, Dauhon Kongeere, No. 29). Herb. Van Royen,
in Rijks Herb., Leiden, sheet "908, 203-…1051" (fide Tanaka,
1932e, p. 422).
Distribution.—India: Coromandel and northern districts to Bhutan; Burma;
Thailand; southern China; North Vietnam; South Vietnam; Laos; Cambodia; Malay
Peninsula; Sumatra; Java; Borneo; Philippines; New Guinea.
Hooker (1875 vol. 1, p. 504) described this
species, with slight rearrangement, as follows: "A [small] tree, branchlets as
thick as a crow-quill and, as well as the inflorescence and leaflets, more or
less softly tomentose; leaves 15-30 foliolate, 6-12 in. [15-30 cm] long;
petiole slender, cylindric, more tomentose than the leaflets, which are very
oblique at the base, petioluled, 2-3.5 in. [5-8 cm] long, ovate or lanceolate,
acuminate, membranous, crenate, upper ones often falcate. Panicle
4-12 in. [10-30 cm] high, pyramidal, branches spreading, alternate.
Flowers 4-merous, shortly pediceled, 1/6 in. [4 mm] diam., white, buds
globose. Petals oblong, glabrous. Ovary ovoid or
elliptic, sub-4-gonal stipitate, hairy or hirsute; style stout, about equaling
the ovary. Fruit 3/4 in. [18 mm] diam., broadly oblong."
Burmann’s original description of C.
excavata (l.c.) covers the stamens, in translation, as follows: "Staminal
filaments 8, subulate, shorter than the corolla, the inferior part dilated and
thickened, hollowed out (‘excavata')2 within."
Guillaumin (1911, p. 661) described the stamens,
in translation, as follows: "Stamens 8, one-third or a half shorter than the
petals; filaments awl-shaped above, then suddenly dilated, concave and
papillose at the base with a hump at the upper part; anthers oval,
apiculate." He also figured the stamens (1911, p. 664, fig.
70 [9-10]), showing the lower expanded and papillose part of the filament to be
strongly curved outward and the upper slender, smooth portion to be even more
strongly curved outward, almost into a semicircle.
The gynoecium of this species, as studied from
serial microtome sections, shows the following structure: pistil 3.2-4.2 mm
long; gynophore very strongly developed, completely glabrous, hourglass-shaped,
usually with a basal annulus 1-1.2 mm diam., then contracted to 0.7-0.8- mm
diam., then expanded into a more or less cupulate top, 1-1.2 mm diam.,
embracing the base of the ovary, with edges often showing small oil glands;
ovary cylindrical or conoid, usually strongly hirsute, 1-5 mm long, 1-1.2 mm
wide, often with a few small oil glands where the style is attached; style
cylindrical, not narrowed at point of attachment, but merging into the top of
the ovary, glabrous, 1-1.2 mm long, 0.5-0.8 mm wide, stigma very short,
flat-topped, only slightly wider than the style. (fig. 3-4,A-B.)
This
species is the type of the genus Clausena and has the widest distribution
of any species, ranging through the East Indian Archipelago to southeastern
Asia. As would be expected from its very wide range, the species
has undergone many variations, until the forms from Sumatra and Taiwan do not
at first sight seem to belong together in a single species.
However, the pistil of this species is distinctive, and is much the same in all
these forms and very different from that of the other species of the genus
Clausena. It has a rather full, smooth, hourglass-shaped
gynophore supporting the more or less strongly hirsute ovary. The
ovary tapers gradually into the thick style, which shows no constriction at its
base as do the styles of many other species.
Only one variety is here recognized, but it would be easy to designate three or
four equally well characterized!
1a. Clausena excavata var. villosa Hook. f. Fl.
Brit. Ind. 1:505. 1875.
Type.—Burma, Rangoon (McLelland, No. 1854). Herb.
Kew.
Distribution.—Upper Burma; also
Thailand, Indo-China, Malaya, and rarely in Nepal (fide Tanaka, 1937, p.
231).
Whole plant clothed with soft spreading
hair: petals hirsute.
This hairy form is found
near the northcentral part of the range of the species.
2. Clausena lansium (Lour.)
Skeels, U.S. Dept. Agri. Bur. Pl. Ind. Bul. 168:3. 1909. Cookia
punctata Sonner. Voy. Ind. Or. 2:231. 1782; Quinaria lansium
Lour. 1790; Cookia wampi Blanco, Fl. Filip. ed. 1. 358. 1837; Clausena
wampi Oliv. Jour. Linn. Soc. Bot. 5(2):34. 1861. Illus.
Sonnerat, loc. cit. pls. 130, 139; Jacquin, Pl. Rar. 1:pl. 101 (col.).
1797; Penzig, Studi Bot. Sugli Agrumi, Atl. pls. 15, 16. 1887; De Wildeman,
Icon. Sel. Hort. Then. 6:pl. 226. 1908; Mill Tsen, Yuan I, Hort.
2:596-97. 1936, 2 text cuts with 6 figs. of fruits and 6 of leaflets; fig.
3-4,E,F,I, and fig. 3-6 this work.
Type.—China, Canton (Loureiro). Herb. Mus. Hist. Nat.,
Paris.
Distribution.—Native in southern
China and Indo-China; widely cultivated in tropical and subtropical
regions.
Common name.—Wampee.
Oliver (1861, p. 34) gave a Latin diagnosis of
this species that, translated, reads as follows: "Tree or shrub; branches at
first pubescent or puberulous; leaves 5-7-9-foliolate, 20-25 cm long; leaflets
ovate-elliptic, lanceolate or ovate, with petiolules 2-6 mm, more or less
oblique at the base, apex obtuse or subemarginate, sometimes almost
mucronulate, slightly acuminate, margin undulate-crenate or slightly serrulate,
becoming glabrous or glabrescent, the midrib and veins often sparsely scabrous,
glabrescent below; terminal leaflet often 6-10 cm long; flowers subsessile or
shortly pedicellate, in many-flowered cymose panicles; calyx-5- (rarely 4-)
merous, lobes triangular or ovate; stamens 10, alternate ones shorter,
filaments dilated above the middle, flattened, subulate above, anthers oblong
or elliptic, cordate-sagittate at base, with a dorsal gland; ovary shortly
stipitate, glandular, very often 5-locular, ovules in pairs [in each locule],
superposed, the upper one peltate on the side or toward the base, the lower one
subpendulous; style very short, distinct, glabrous above, stigma 5-lobed,
slightly wider than style; fruit usually with 5 locules, 5-seeded, or by
abortion 1-seeded, or sterile, ovoid-globose, pubescent, 1 in. [25 mm] or less
diam.; cotyledons fleshy, equal."
This
species differs widely from all the other species of the genus, as has been noted. The wampee is a highly esteemed fruit tree in
southern China, where sour, subacid, and sweet varieties are known.
The white or yellow fruits are ovoid or subglobose, about the size of a
pigeon’s egg, but varying in size and shape with the variety
cultivated. The inflorescences are large panicles at the ends of
the branches, so the fruits occur in rather openly-spaced clusters.
In texture, the wampee fruit is much like a loquat and is a berry without a
tough peel. An illustrated treatise in Chinese by Mill Tsen [Ts’êng
Mien-chih, in Mandarin transliteration] (1936) described in detail six
varieties and figured the fruit and a leaflet of each variety.
(Mill Tsen mentioned another variety [no. 6 below], which is not
figured.) Table 3-1 shows the principal characters of the fruits of these
Foochow varieties (the leaves vary almost as much as the fruits but are omitted
from the table).
According to Mill Tsen,
Professor Wên Wên-kuang of Sun Yat-sen University at Canton, China, listed
eight varieties of the wampee grown in Kwangtung Province. These
varieties are not described in detail but are given long descriptive names
such as "white-hairy-chicken-heart-sweet-wampee";
"long-chicken-heart-sour-wampee"; "yellow-hairy-chicken-heart-sour-wampee,"
etc.
Although the wampee is only remotely related to Citrus, it
can be grafted on a Citrus rootstock, and is thereby forced into early
flowering and fruiting. The rough lemon can be grafted on the
wampee and will grow for many years if a small branch of the wampee is allowed
to grow just below the graft. Any desired Citrus fruit can
be top-grafted on the rough lemon and may live for many years and may even be
made to fruit. However, the graft union of the rough lemon on the
wampee is poor and badly overgrown. Wampee on Citrus seems
to make a much better and more permanent union.
3. Clausena pentaphylla
(Roxb.) DC. Prodr. 1:538. 1824. Amyris pentaphylla Roxb. Fl. Ind.
2:247. 1832. Illus. fig. 3-4,C.
Type.—British India, United Provinces, Cawnpore, a living plant sent to
Calcutta Bot. Gard. and grown there (Roxburgh). Herb. Brit. Mus.,
London.
Distribution.—India: western
Himalayan region, from Garhwal to Sikkim, northern Uttar Pradesh and Oudh.
Brandis (1906, p. 114) gave a condensed
description of the species as follows: "A strongly aromatic shrub, young shoots
and inflorescences silky-tomentose, full-grown leaves pubescent.
Leaflets 5 or 7, nearly opposite, 4-6 in. [10-15 cm] long, secondary nerves
prominent beneath. Flowers yellowish, in terminal
panicles. Berry verrucose, pale orange, 1/3-1/2 in. [8-13 mm]
long." Duthie (1905, p. 138) in his Flora of the Upper Ganghetic
Plain, a region where this is the only native species of Clausena,
described it as follows: "A deciduous shrub; young parts
silky-tomentose. Leaves large, 3-7-foliolate. Leaflets
subopposite or alternate, 2-6 in. [5-15 cm] long, ovate or lanceolate,
acuminate, entire, shortly stalked, softly tomentose beneath; main lateral
nerves prominent. Flowers 4-merous, 1/4 in. [6 mm] across,
yellowish, in erect terminal downy panicles. Sepals triangular,
acute. Petals oblong, concave. Filaments short,
broad. Berry 1/2 in. [13 mm] diam., ovoid, papillose, pale
orange."
Good flowering material collected by
J. F. Duthie in the western Himalayan region at Gonda Dun (No. 21613, Herb.
Arnold Arbor.) shows the following gynoecium characters: The pistil, 2-2.2 mm
long, shows no sharp boundaries of disk, ovary, style, and stigma.
The 3- or 4-locular ovary is 1-1.2 mm wide and 0.9-1 mm long, irregularly
tuberculate and sparingly pilose; the style is cylindrical above, 0.35-0.45 mm
wide, with no obvious stigma; below, it may be slightly contracted where it
joins the top of the ovary, but very often it is swollen with rather large oil
glands at or just above its junction with the tip of the ovary and in
consequence may be nearly as wide as the ovary. The disk is most
irregular, usually as broad as the ovary but more or less lobed and even having
short processes each ending in an oil gland. The locules of the
ovary run deeply into the disk, which in longitudinal section is not clearly
separated from the base of the ovary.
This
species, remarkable for its deciduous foliage and shrubby habit, has even more
remarkable pistils that depart widely in their characters from those of all
other species of the genus. Its nearest analogue in gynophore
morphology is the anomalous C. lenis, which has a more or less syncopated gynoecium.
4. Clausena indica (Dalz.)
Oliv. Jour. Linn. Soc. Bot. 5(2):36. 1861. Piptostylis indica
Dalz. Hook. Jour. Bot. & Kew Gard. Misc. 3:33. 1851; Bergera
nitida Thw. Enum. Pl. Zeyl. 46. 1858. Illus. Dalzell, loc. cit.
pl. 2; Engler, Die Nat. Pflanzenfam. 3(4):187, fig. 108,L-O.
1896; ibid. ed. 2. 19a:321, fig. 146,L-O. 1931; Talbot,
Forest Fl. Bomb. Presid. 1:195, fig. 119. 1909; fig. 3-4,K this work.
Type.—British India, Bombay Ghats, Kanara (Dalzell). Herb.
Kew.
Distribution.—India: western
peninsula, Bombay Ghats to Anaimalai Hills, Madras State, eastern coast,
Tranquebar; Ceylon.
Hooker (1875, vol. 1, p.
505) gave a description that may be summarized as follows: "A glabrous shrub or
small tree; leaves 9-11-foliolate, 4-10 in. [10-25 cm] long; petioles
slender…glabrous or puberulous; leaflets polymorphous, very oblique
oblong-ovate, elliptic or lanceolate, rarely almost rounded, tip
rounded-obtuse and notched or acute or acuminate, margins crenulate, very dark
with raised glands on both sides when dry; inflorescence a peduncled panicle,
corymbosely branched above; flowers 1/6 in. [4 mm] diam., very shortly
pediceled; ovary 2-5 celled, glabrous, papillose; ovules 2 in each cell; fruit
1/2 in. [13 mm] diam., globose, yellow, edible." Talbot (1909)
added a few details that can be condensed as follows: Leaflets subcoriaceous,
2-4 X 1-2 in. [5-10 X 2.5-5 cm]; petiolules 1/4 in. [6 mm] long; petals broadly
ovate; stamens with filaments broad, as long or longer than the versatile
anthers; ovary usually 3-loculed; fruit pulpy, edible. Dalzell
(l.c.) described and figured the style as much shorter than the ovary, thick,
cylindrical, with the stigma scarcely wider than the style, which is not
expanded at the base where it is situated in a depression at the tip of the
ovary and wholly caducous. He described the flowers as 4- or
5-merous but did not see any but 3-locular
ovaries.
Clausena indica is a shrub or
small tree, 2 to 2.3 m high, common in the evergreen rain forests of the Ghats
of western peninsular India. In spite of its rather limited
distribution, it shows great variation in the shape of the leaflets which
differ greatly in outline, being "very oblique" in Ceylon, according to Trimen
(1893, p. 221), but only slightly if at all oblique at the base in the type
locality in the Bombay Ghats, as is shown by Dalzell's careful drawing (and
even more by his type specimen in the Kew herbarium) as well as by Talbot's
sketch. The ovary varies even more strikingly than the leaflets,
having regularly three locules in the type locality, according to Dalzell
(l. c.) and Talbot (1909), whereas in Ceylon it always has two locules,
fide Dalzell (l. c.) and Trimen (1893, p. 221). A
specimen from Ceylon sent by the Royal Botanic Garden at Peradeniya to the
herbarium of the Arnold Arboretum (Tanaka's "Det. A-229") has 2-, 3-, and
4-locular ovaries in the same inflorescence. Oliver and Hooker both
found forms with 4- or 5-locular ovaries.
As
this species bears edible fruits, it should be hybridized, if possible, with
the Chinese wampee and with other species that yield edible fruits.
5. Clausena heptaphylla
(Roxb.) Wt. & Arn. Prodr. 96. 1834; and in Wall. Cat. 8508. 1848. Amyris
heptaphylla Roxb. Fl. Ind. 2:248. 1832; Clausena forbesii
Engl. 1896. Illus. Guillaumin, in Lecomte, Fl. Gén. Indo-Chine
1:664, fig. 70(11-12). 1911; Tillson & Bamford, Amer. Jour. Bot.
25:782, fig. 14. 1938; fig. 3-4,D this work.
Type.—British India, "about Calcutta" (Roxburgh). Herb. Brit.
Mus., London.
Distribution.—India,
Burma, Thailand, North Vietnam, South Vietnam, Laos, Cambodia and Sumatra (also
Java, fide Tanaka).
The description of
this species given by Guillaumin (1911, p. 662) reads, translated, as follows:
"A shrub; leaves 30-50 cm long; leaflets 7-11, unequal-sided at base (except
the terminal one), membranaceous, rigid, long-oval or lanceolate, 4.5-16 X
2.5-7 cm, acute at base, acuminate at apex, entire, glabrous except the midrib
on the under surface, with 6-7 pairs of lateral veins which are raised on the
under surface; petioles cylindrical, with very short pubescence,
glandular. Inflorescences terminal, paniculate, pyramidal, shorter
than the leaves, with alternate, slightly and shortly pubescent branches;
pedicels very short with 2 basal, shortly pubescent bracts articulated at the
very base. Sepals 4, triangular, 1 mm long with ciliate
margins. Petals 4, obovate, glabrous but papillose, twice as long
as the sepals. Stamens 10, slightly shorter than the petals;
filaments filiform at base then abruptly dilated below the anther; anthers
attached dorsally along the middle, almost rhombohedral, without
horns. Disk very short, glabrous. Ovary cylindric,
quadrangular glabrous, as long as the stamens, 4-locular, each with 2
superimposed ovules; style cylindrical, as long as the ovary. Fruit
ovoid, 2 X 1 cm, very glandular, with 1 or 2 seeds."
Hooker (1875, vol. 1, p. 504), who described the
species from material from a wide range of localities, noted that it is a
"branching bush, smelling strongly of aniseed.…Leaves 6-16 in. [15-40 cm
long]…leaflets sometimes 8 in. [20 cm long]…membranous, ovate or lanceolate,
acuminate, very obscurely crenulate, pale beneath, nerves and costa
slender.…Flowers 1/6 in. [6 mm] diam., pedicels slender, buds glabrous,
yellow-white.…Berry 1/2 in. [12 mm] long, oblong, white-reddish or pale yellow,
glandular."
Engler (1931, p. 321) mentioned
in his notes on Clausena: "C. forbesii [from Sumatra] with leaves
nearly one meter long, having eight paired leaflets," the longest leaves yet
reported in the genus. Probably it is merely an unusually vigorous
growing form of this polymorphic species.
This species, in spite of its abundance, needs renewed study to determine its
limits of distribution in the East Indian Archipelago. Tanaka
(1937, p. 231) even reported it from the Sunda Islands and Timor.
It has not been reported from Ceylon or from Borneo or the Philippines.
5a. Clausena
heptaphylla var. engleri (Tan.) Swing. Jour. Wash. Acad. Sci.
32:26. 1942. Clausena engleri Tan. Med. Rijks Herb. Leiden
69:6. 1931; (?) C. platyphylla Merr. Pap. Mich. Acad. Sci.
23:182. 1938.
Type.—Sumatra
(Lörzing, No. 6825). Rijks Herb.,
Leiden.
Distribution.—Known only from
Sumatra.
Tanaka's original description reads,
translated, as follows: "A robust plant; rachis of leaf 36 cm long,
7-foliolate; leaflets ovate, acuminate-caudate, pointed at the base, with
conspicuous veins, when dry chartaceous, very green on the upper surface,
pallid below, margins serrate, veins rufescent, with very short petiolules,
entirely glabrous. Inflorescence terminal, 28 cm long, branched
into many-flowered panicles. Flowers small, 5-merous,
short-pedicellate. Calyx thin, scutellate, with triangular pilose
lobes having ciliate margins. Petals 5, caducous.
Stamens 10, filaments dilated, anthers large. Ovary cylindrical,
pilose; style linear, straight or curved, stigma somewhat capitate."
Tanaka, in his remarks following the Latin
description, stated that this plant resembles the robust form of C.
heptaphylla, but the latter has 4-merous flowers, glabrous ovary, and entire
leaflets." The photograph of Tanaka's type specimen shows it to
have large leaflets, 11 to 19 by 5 to 8 cm, at the apex, cuneate and only
slightly unequal-sided at the base, with 10 to 12 pairs of lateral
veins. C. platyphylla, recently described as a new species
from Asahan in northeastern Sumatra, seems likely to prove to be a synonym of
C. heptaphylla var. engleri once a comparative study is made of
the type specimens of these two plants. Both of them have leaflets
of about the same size, of similar shape, with the same number of lateral
veins, and even share the peculiarity of the dried leaves being dark, shining
green above and much lighter, pallid green below. The ovary of var.
engleri is pilose, whereas that of C. platyphylla shows scattered
pubescence; the ovary of the species C. heptaphylla is glabrous.
6. Clausena dentata (Willd.)
Roem. Syn. Hesper. 44. 1846. Amyris dentata Willd. Sp. Pl. 2:337.
1799, Clausena willdenowii Wt. & Arn. 1834. Illus.
Wight, Icon. Pl. Ind. Or. 1:pl. 14/339. 1838; Beddome, Fl. Sylv. Anal.
Gen. pl. 7. 1871.
Type.—British India
(Willdenow, No. 7293). Herb. Bot. Mus.,
Berlin-Dahlem.
Distribution.—Ceylon;
India: western peninsula, eastern Himalayan region, Sikkim to northwestern
Burma; Thailand; southwestern China.
This
species was described by Hooker fils (1875, vol. 1, p. 506), with some
transpositions, as follows: "A large shrub, glabrous or…pubescent; branchlets
slender. Leaves 5-13-foliolate, 6-16 in. [15-40 cm] long; petiole
slender, terete; leaflets 1-2 in. [2.5-5 cm] long, oblique, oblong-ovate or
ovate-lanceolate, obtuse, acute or obtusely caudate-acuminate, crenulate,
membranous. Racemes slender, axillary, 3-10 in. [7.5-25 cm] long,
branches or pedicels divaricating. Flowers fragrant, usually
4-merous, 1/3 in. [8 mm] diam., whitish; buds globose. Petals
oblong, concave, glandular, ovary stipitate, 4-angled or grooved, glabrous,
ovules 2, superimposed in each cell; style short, thick. Fruit
globose, from the size of a pea to a cherry, whitish-green, pellucid."
A specimen from southern India, Madras State,
Hosur Taluk (coll. by Mrs. Kanoth Yeshoda; now in Herb. Arnold Arbor.), shows
the restored flowers in serial microtome sections to have the following
characters: pistil, 3.5 mm long; gynophore short, 0.5-0.6 mm long, narrowed to
0.7 mm just below the flatly rounded, nearly truncate ovary base; ovary
subglobose, 1.3-1.4 mm long, 1.2-1.3 mm wide, tip abruptly rounded with a slight
depression in which the nearly cylindrical style is attached; style 1.5 mm
long, 0.3-0.4 mm wide, stigma short, cushion-like, 0.5 mm wide.
Cross sections show 4-locular ovary with a very short process at the tip of each
locule but with no large oil gland. The pedicels of the flowers are
unusually long (4-6 mm) and slender, with small bracts at the base.
This species, like C. suffruticosa and the
three African species, C. abyssinica, C. anisata, and C.
inaequalis, has axillary flower clusters, not terminal panicles as in most
of the other species. The long pedicels of the flowers are unusual
in Clausena.
6a.
Clausena dentata var. pubescens (Wt. & Arn.) Tan. Jour. Bot.
Brit & For. 68:227. 1930. Clausena pubescens Wt. &
Arn. Prodr. 1: 96. 1834; C. willdenowii var. pubescens (Wt.
& Arn.) Hook. f. Fl. Brit. Ind. 1:506.
1875.
Type.—Ceylon (Wight, No.
328b). Herb. Glasgow
Univ.
Distribution.—Ceylon; India:
Mysore State.
Tanaka (1930b, p. 227)
described this variety, in translation, as follows: "Leaves, inflorescences,
and petioles densely villose. Fruits globose, rather small."
Tanaka (1937, p. 230) stated that this variety is
very rare in Mysore and Ceylon.
6b. Clausena dentata var. longipes (Craib) Tan.
Bul. Soc. Bot. France 75:709. 1928. Clausena longipes Craib, Kew.
Bul. Misc. Inform. 1926:340. 1926.
Type.—Mê Lan, Mê Hawng Sawa, Thailand, 700 m altitude, on limestone rock
(Kerr, No. 5486-A). Herb. Kew.
Distribution.—Thailand and upper Burma.
Differs from the species in having (1) very slender, few-flowered
inflorescences ending in long, slender pedicels, 5-15 X 0.2-0.3 mm; (2) slender
(1.7-2-5 X 1 mm) ovaries that merge gradually into the rather long style (2.5-3
X 0.6-0.7 mm); (3) no oil gland at the tip of each of the 4 locules of the
ovary; (4) very narrow radial locule walls, not swollen where they cross to
make an axial pillar; and (5) few tannin cells in the periphery of the ovary
and none in the axis.
This variety appears at
first sight to be specifically distinct from C. dentata, but some of the
material of the species from southern India shows very slender inflorescences
and almost equal-sided leaflets very like those of var.
longipes. Craib (1926, p. 234) considered this plant to be
related to C. dentata and to C. suffruticosa.
6c. Clausena dentata
var. dulcis (Bedd.) Swing. Jour. Wash. Acad. Sci. 28:532. 1938.
Cookia dulcis Bedd. Madras Jour. Lit. Sci. 2 ser. 22:73. 1861.
Clausena willdenowii var. dulcis Bedd. Fl. Sylv. 44.
1871.
Type.—British India, Madras
Presidency, Anaimalai Hills (Beddome). Herb.
Kew.
Distribution.—India: Anaimalai
Hills south to Travancore.
Common
name.—Indian wampee.
Beddome (1861, p. 73)
described his new species in general terms as follows: "A tree with a delicious
fruit not uncommon on the Anamallays up to 3,000 feet both in the moist woods
and in the drier forests—it flowers in April and the fruit begins to ripen at
the end of June—the fruit is more grateful to the taste than that of the
Whampee (Cookia punctata). The tree is well known to the hill
tribes and called 'Mor Koorangee.’ I have often met Kaders carrying
home on their backs basket loads of this and the fruit of Pierardia
sapida, which is also abundant in these jungles." He also noted
that the "fruits are globose, size of a large cherry." Beddome in a
later publication (1871, p. 44) stated: "This is a tree 30 feet high, with a
trunk several feet in girth, bearing a very delicious fruit as large as a large
cherry, as succulent as a grape, and somewhat of the flavour of the black
current…it is abundant about Toonacodava (elevation 2,000-3,000 ft.), both in
the moist and dry forests, and is in fruit from June till August; specimens in
flower were sent to Prof. Oliver, who pronounces it only a form of
[Clausena] Willdenowii [= C. dentata], so I will not
attempt to separate it, but I believe it quite a distinct species…in comparison
with true C. Willdenowii it has leaves that are more membranaceous,
highly odoriferous, more prominently dotted, and very erose toward the apex."
This variety is not as yet clearly
distinguished from the species, and most taxonomists have not recognized it as
valid. It should be noted, however, that investigations by Rao and
Subramaniam (1935) have shown that the two forms of C. willdenowii [=
C. dentata] growing in southern peninsular India (Madras State) differ
strikingly in the composition of the volatile oil in the leaves. Oil
from the leaves of one form was shown to consist of alpha-,
beta-, and di-alpha-clausenan belonging to the furfural group, of
which the major constituent was alpha-clausenan. Oil from
leaves from a different locality was found to contain not alpha-clausenan
but gamma-clausenan, an isomeric substance.
Abundant good herbarium specimens from Madras
State, lent to Swingle with other material of Clausena by the Director of
the Royal Botanic Gardens at Kew, show a decided variation in the shape of the
ovary and in the relative length of the style. A specimen collected
by J. S. Gamble (No. 16420, July, 1885, the Chingleput district of Madras
State) shows abundant flowers with a slender, often conical ovary, 1 to 1.2 mm
long and 0.6 to 0.7 mm wide, merging into a cylindrical style distinctly longer,
1.2 to 1.8 mm (all measured dry). Another specimen collected by
Gamble (No. 16190, Barliyár, Nilgiris, at alt. 3,000 ft. [915 m], May, 1885)
shows equally abundant flowers with a more or less cubical, 4-angled ovary
about 1 by 1 by 1 mm with a prominent oil gland at the tip of each of the four
locules. The cylindrical style does not merge into the tip of the
ovary but is inserted between the four prominent oil glands; it is only
slightly longer than the ovary, 1.1 to 1.2 mm when dry. Another
specimen, Wight's No. 354 (collected at Courtallem [Kattalan], July, 1835), has
more or less cubical, 4-angled ovaries with the style distinctly longer, about
1.5 mm when dry.
Evidently careful research
is needed to distinguish the forms of this and possibly other varieties from
the species. As var. dulcis is of economic importance, it is
to be hoped that its taxonomic status can be settled soon.
Other observers agree with Beddome concerning
the delicious character of the fruit of this variety growing in the western
part of Madras State. Thomas A. Bourdillon (1908, pp. 67, 68), for
more than thirty years Conservator of Forests in the former native state of
Travancore, stated: "I agree with Beddome that the fruit, which very much
resembles a grape, is very delicious, and is well worthy of
attention." He found it "common in the evergreen forests of
Travancore at an altitude of 1,000-4,000 ft. [305-1,220 m] where the rainfall
is light. It is a small tree, 30 ft. [9.14 m] high and 9 in. [23
cm] in diameter, that is leafless in March, flowers in April and fruits in May
and June."
J. S. Gamble, in his Flora of
the Presidency of Madras (1915, p. 155), referring to C. willdenowi
[= C. dentata], probably in large part this var. dulcis, stated:
"A small tree with glabrous or pubescent leaves, black bark and white,
close-grained wood. The fruit is good and worth cultivation!"
6d. Clausena dentata
var. dunniana (Lév.) Swing. Jour. Wash. Acad. Sci. 30:82 1940.
Clausena dunniana Lév. in Fedde, Repert Sp. Nov. 11:67.
1911.
Type.—China, Kweichow Province,
Pin-fa (Cavalerie, No. 1072). Herb. Univ.
Edinburgh.
Distribution.—China:
Kweichow Province.
Leaves 5-11-foliolate;
leaflets elliptical, bluntly pointed at apex, 5-10 X 2.5-3.5 cm, very blunt
(almost truncate) at base, more or less unequal-sided, inconspicuously
crenulate; petiolules unusually long (3-7 mm); inflorescences axillary,
slender; flower buds globose, small, 2 mm diam.; petals glabrous; pistil 2.5 mm
long, glabrous; gynophore 0.8 mm long, 0.6 mm wide; ovary tuberculate, 1 X 1
cm, with 1 or 2 small or medium-sized oil glands in a tubercule that tips each
locule; style cylindric, 0.8-0.9 X 0.5 mm, narrowed at base; fruits subglobose
or short-ovoid, 7-8 X 6-7 mm, black.
Little is
known about this shrubby variety that grows at altitudes of from 200 to 1,200
m. It may prove to be a good species as yet inadequately
described. The oil glands in the apical tubercles of each locule
of the ovary are somewhat like those of C. dentata, although usually
only one medium-sized oil gland is found in the species. For the
present we are considering this Chinese plant as a variety of C.
dentata, which also has axillary inflorescences.
6e. Clausena dentata
var. robusta Tan. Jour. Bot. Brit. & For. 68:228.
1930.
Type.—China, Yunnan Province,
Szemao; altitude 1,372 m (Henry, No. 13028). Herb.
Kew.
Distribution.—China: Yunnan
Province.
Tanaka's original description reads,
in translation, as follows: "A vigorous plant, leaflets large, broadly oval,
unequal-sided rather thick. Inflorescences suberect,
robust. Berry globose, pustulate-punctate."
Tanaka cited two numbers as types: Henry, Nos.
11914D and 13032, both in the herbarium at Kew. The senior
author could find only No. 11914D, which consists of the tip of a
fruiting branch with a single leaf and 4 axillary inflorescences, 12-18 X
3.5-4.5 cm. The leaflets are 8.5-9 X 3-3.8 cm and are acuminate;
the berries are subglobose, 7-8 mm diam., borne on a twig 5-7 mm diam., with
elongate heart-shaped leaf scars, 6 X 3 mm. Another sheet in the
Kew Herbarium is Henry's No. 13028, also from Szemao, with flowering twigs with
11 foliolate leaves, 32-37 X 12-14 cm; lateral leaflets 5.5-9 X 3.5-4.5 cm,
terminal 10 X 5 cm; inflorescences 12-14 X 2.5-3 cm; flower buds globose or
obovoid, 2.5 mm long, 2.5-3 mm wide; pistil glabrous, 3 mm long; gynophore
0.7-0.8 X 0.5 mm; ovary tuberculate, 0.7-0.8 mm wide; style cylindric, 1.5 X
0.6 mm, stigma depressed, slightly wider than the style. The
leaflets are widely spaced on the rachis (2-4 cm) like those of C. dentata
var. dunniana, from which var. robusta is not adequately
separated by the brief original description.
6f. Clausena dentata var. henryi Swing. Jour.
Wash. Acad. Sci. 30:81. 1940.
Type.—China, Hupeh Province, Ichang (Henry, No. 4122).
Cotype: Ichang (Henry, No. 3127). Both in Herb. Kew.
Distribution.—China: known only from
western Hupeh and Yunnan (?) provinces.
Differs from C. dentata var. robusta Tan., from Yunnan, in having
hirsute petals, ovate black fruits, and more densely hirsute young
leaves. A small tree, 2-6 m high; leaves persistent, 25-35 cm long,
13-17-foliolate; leaflets lanceolate, acuminate at tip, blunt at base, which is
often strong (and always somewhat) unequal-sided, 5-12 X 2-4.5 cm; largest
leaflets often laterals, attached near upper third of rachis; rachis, midribs,
veins, and whole surface of young leaflets pubescent, but leaflets glabrescent
above; inflorescences axillary, sometimes short, 8-10 cm, often longer, 20-25
X 4-6 cm, pubescent when young but soon glabrescent; fruits black when ripe,
ovoid, 9-12 X 6-8 mm.
The fruits of var.
henryi are considered edible in central China, according to Rehder and
Wilson (1914, p. 140). It has been thought by some botanists that
this plant is sometimes cultivated in Hupeh Province. It should be
introduced into culture in other regions and hybridized with species that
produce edible fruits.
7. Clausena harmandiana (Pierre) Guill. in Lecomte, Not.
Syst. 1:219. 1910. Glycosmis harmandiana Pierre, Flore Forest.
Cochinch. 4(18):text to pl. 285. 1893. Illus. Guillaumin, in
Lecomte, Fl. Gén. Indo-Chine 1:664, fig. 70(4-5).
1911.
Type.—French Indo-China, Cambodia
(Harmand, No. 3875). Herb. Mus. Hist. Nat.,
Paris.
Distribution.—Cambodia and
Laos. According to Tanaka (1937), it was collected once at Margui,
Burma.
The original description by Pierre
reads, translated, as follows: "Branches rounded, pubescent. Leaves
imparipinnate, with 6-9 opposite or alternate leaflets, which are subdeltoid,
ovate-acuminate, obtuse at both ends, 5-14 cm long, 5-8 cm wide, submembranous,
coriaceous, with 7-8 pairs of small lateral veins. Inflorescences
terminal, pubescent, 10-20 cm long, much branched with rather large (5-6 mm
diam.), sessile flowers. Sepals 1 mm long, very coriaceous,
pubescent on the outside and ciliate. Petals 3.5-4 mm long, very
concave, obovate, veinless, red spotted. Stamens 7-10, 2.5 mm long,
with the filaments flattened below and subulate above, longer than the anther,
which is oval and without an apical gland. Disk
cupuliform. Ovary 2.5 mm long, with 5 locules, each carrying on the
outside 2 large glands. Ovules 2 in each locule."
The description of the disk by Pierre as
cupulate is difficult to credit. A photograph of the flowering
type specimen (Harman, No. 3875) in the Paris herbarium, placed at Swingle's
disposal by Tanaka, led him to think that Pierre mistook a ring of small bracts
at the base of the very short pedicel (only about 1 mm long) for the calyx and
the calyx itself for the disk! The length of the flower from the
base of the calyx to the stigma measures about 3 mm, the calyx flattened-rotate,
about 2 to 2.5 mm wide, tapering rapidly into a very short pedicel about 1 mm
long (including the bluntly conical base of the calyx); at the base of the
pedicel are several small bracts and sometimes one or two small flower buds
which may develop later, or more often abort and fall off. These
bracts and buds are borne on a slender terminal branchlet of the inflorescence
3.5 to 8 mm long. The ovary is subglobose or broadly oval, about
1.2 to 1.5 mm long and 1 mm wide; the style is cylindrical, 0.4 to 0.5 mm long
and 0.2 to 0.3 mm wide, not contracted at the base where it joins the ovary
tip. The gynophore is shorter than in most species of
Clausena, but seems to be otherwise normal.
7a. Clausena
harmandiana var. contracta Tan. Med. Rijks Herb. Leiden 69:7.
1931.
Type.—Java
(Horsfield). Herb. Kew.
Distribution.—Known only from Java.
The
type specimen of this variety has 5-7 leaflets varying in size, the lower ones
3-4 X 2-2.5 cm, the larger laterals 7-9 X 3-3.5 cm and the terminal one 10 X 4
cm, all nearly equal-sided, broad-elliptical, blunt at both ends, all parts
pubescent, even the upper surface of the leaflets slightly so; inflorescences
terminal, very short, 8 X 3 cm; flower buds globose, 2 mm diam., yellowish to
reddish-brown when dry; petals without large oil glands at tips; gynophore 1 mm
long; ovary 1 mm long, 1.5 mm wide, jet black, glabrous; style 0.5 mm long,
glabrous, black.
Another specimen in the
herbarium at Kew, identified by Tanaka as this variety (coll. by Zollinger,
second Java voyage, No. 2878), has more glabrous leaves; ovary jet black, very
prominently tuberculate; gynophore subcylindric, abruptly expanding into the
ovary base; style 0.6 mm long, 0.5 mm wide, stigma depressed-capitate, 0.7 mm
wide.
This variety is apparently known only
from these two specimens and is not well understood. Tanaka
(1931b) stated that it resembles somewhat C. dentata var.
robusta of China, but differs in having terminal, not axillary, short
inflorescences and in having a shorter style and a thicker ovary.
8. Clausena macrophylla Hook.
f., Fl. Brit. Ind. 1:504. 1875. Clausena heptaphylla var. (?)
pubescens Oliv. Jour. Linn. Soc. Bot. 5(2):30. 1861; C. hirta
Ridl., pro parte (?) 1920. Illus. fig. 3-4,G.
Type.—Burma, Trogla, banks of Saluen River (Wallich, No. 6367).
Herb. Kew.
Distribution.—Burma;
Thailand: Phuket, Tasan; Malay Peninsula.
Hooker (1875, vol. 1, p. 504) described this species, with some transpositions
as follows: "Branchlets as thick as a goose quill and, as well as the petiole
and panicle, clothed with a soft, spreading, close-set velvety
tomentum. Leaves 5-foliolate, 1 ft. [30 cm] long, leaflets 5-7 in.
[12.5-17.5 cm] long, equal or suboblique with very short petiolules, acuminate,
obscurely toothed, small, rather distant teeth, pale on both surfaces,
tomentose beneath, nerves strongly arched, midrib stout, common petiole stout,
terete. Panicle stout, erect, with the peduncle 1 ft. [30 cm]
long; branches strict, erecto-patent, secondary branches short.
Flowers glabrous, crowded, 1/10 in. [2.5 mm] diam., shortly pediceled, buds
globose. Calyx lobes 4, minute, rounded-acute. Petals
broadly oblong, concave, membranous, covered with large glands.
Stamens equal; filaments much dilated below, very short; anthers large,
oblong. Ovary stipitate, glabrous, 4-lobed, tuberculed with very
large glands; ovules 2, collateral in each cell; style as long as the ovary and
as broad as the stigma. Fruit immature, oblong, tip obscurely
4-angled and truncate."
Ridley mentioned in
his description of C. hirta that the leaflets are "thickly covered with
translucent glands and dotted over with larger, sparser dark brown
ones." This description recalls the dimorphic oil glands of C.
guillauminii and makes it very desirable to reexamine all material of C.
hirta to see if any confusion has arisen with C. guillauminii, a
species which also occurs in Thailand.
9. Clausena anisum-olens (Bl.) Merr. Phil. Gov. Lab. Bur.
Bul. 17:21. 1904. Cookia anisum-olens Blanco, Fl. Filip. 359.
1837; Clausena warburgii Perk. Frag. Fl. Phil. 162. 1905; C.
loheri Merr. Phil. Jour. Sci. 27:27.
1925.
Type.—Lacking.
Substitute type: Rizal Province, Luzon, P.I. (Merrill, Species
Blancoanae, No. 1012). Herb. Bur. Sci.,
Manila.
Distribution.—Philippine
Islands: Luzon, Masbate, Basilan, Mindanao islands.
Perkins (l.c.) has drawn up the only full
description of this species yet published. It reads, translated, as
follows: "Shrub or tree, with glabrous…twigs 5 mm thick; leaves 15-30 cm long,
2-paired…very sparsely gray-pilose; leaflets ovate, or obovate-oblong or
ovate-lanceolate, 6.5-10 cm long, 2.5-3 cm wide, apex long, narrowly acuminate,
the apex itself obtuse or often subincised, base oblique, papyraceous,
glabrous, veins and veinlets sparsely pilose…lateral veins 8-12 [pairs], veins
and veinlets indistinct above, prominent below. Inflorescences
terminal, paniculate, 18-22 cm long, branchlets numerous, sparsely pilose,
spreading, bractlets ovate, sparsely pilose; flowers 8 mm diam., with pedicels
5-8 mm long; calyx 5-merous with short lobes…ovate-acuminate, very sparsely
pilose without; 1 mm long; petals with lobes…5 mm long, 2 mm wide, ovate,
membranaceous, glabrous on both sides; stamens 9, as long as the petals,
filaments jointed (geniculate) near the middle, the joint pubescent; disk
small, smooth, ovary stipitate, glabrous, 5-locular, 1 mm diam.; style as long
as the ovary, stigma sessile, 5-lobed."
Serial microtome sections of the pistil of a specimen in the Bureau of Science
Herbarium at Manila (coll. by E. D. Merrill, No. 2509, Bur. of Agric., at Lamao
River) show the following characters: Pistils 5-locular, each locule ending in a
short process, bearing at the end a rather large oil gland; ovary broad-oval,
about 1.5 mm long, 1-1.2 mm wide; style strongly contracted at base, which is
inserted into a cavity at the tip of the ovary, the rest of the style
unusually broad, nearly 0.8-1 mm wide; the gynophore is long and not abruptly
narrowed, about 1-1.2 mm long, 0.7-0.8 mm wide at the narrowest place, and about
1 mm at the base and the top.
This species is
confined to the main islands of the Philippines. It has geniculate
staminal filaments, bent at an angle near the middle, slender above the bend
and broader below, resembling somewhat the filaments of C. excavata, but
it is much less bent and much less swollen below the bend.
It is related to C. laxiflora of Mindanao
Island, Philippines, but shows important differences in the characters of the
pistil and the inflorescences.
10.
Clausena mollis Merr. Phil. Jour. Sci. Bot. 5:181.
1910.
Type.—Philippines, Luzon Island,
near Bontoc, at 915-1,220 m attitude (Curran, For. Bur., No.
16530). Herb. Bur. Sci.,
Manila.
Distribution.—Philippines:
known only from Luzon Island.
Merrill's
original short diagnosis, translated and supplemented with data from the longer
description (in brackets), reads as follows: "A shrub about 5 m high, all parts
densely and softly pubescent; leaves 20-30 cm long; leaflets alternate or
subalternate, 5-8 on each side, margins entire, base strongly inequilateral,
apex shortly acuminate or…blunt, [lateral veins about 8 on each side of the
midrib]. Panicles subterminal, narrowly pyramidal, flowers
5-merous, sessile or subsessile, subglomerate [on the ultimate branches]…Sepals
broadly ovate, 1.2 mm long, free, densely pubescent outside. Petals
elliptic or broadly elliptic-oblong, concave, about 4 mm long, 2.5 mm wide,
acute at both ends, imbricate, with few, rather large glands, the back in the
upper third slightly pubescent. Stamens 10, the filaments broad,
abruptly narrowed just below the insertion of the anthers, about 1 mm long,
anthers 2 mm long. Ovary ovoid or ellipsoid, about 1.5 mm long,
glabrous, prominently 5-sulcate, 5-celled, each cell with two superimposed
ovules; style 1 mm long and anthers 2 mm long. Ovary ovoid or
ellipsoid, about 1.5 mm long, glabrous, prominently 5-sulcate, 5-celled, each
cell with two superimposed ovules; style 1 mm long and thick
[5-sulcate]. Fruits globose, white or greenish-white,
1-seeded. [Seeds about 5 mm diam., surrounded by a gelatinous pulp,
with a strong odor and taste of pine pitch.]"
This species is doubtless related to C. anisum-olens but differs from it,
not only in being densely soft-woolly all over (except the petals and the
flower parts they enclose), but also in having pistils with a rather short
gynophore (somewhat like that of C. laxiflora). The filaments
of the stamens are abruptly expanded from the middle down, and somewhat
geniculate where the slender apical portion joins the expanded base.
Each locule of the ovary is capped by a single large oil gland. The
flowering twigs of this species are often unusually thick; a specimen collected
by Ramos and Edaño (Bur. Sci., No. 7876), at Mount Masapilid, Bontoc Province,
Luzon Island, in March, 1920, now in the herbarium of the Arnold Arboretum, has
twigs 5 to 7 mm in diameter, with reddish-brown pith 2.5 to 3 mm in
diameter.
11. Clausena
laxiflora Quis. & Merr. Phil. Jour. Sci. 37:154. 1928.
Illus. fig. 3-4,M.
Type.—Philippines, Mindanao Island, Davao Province, Mati (Ramos and Edaño,
Bur. Sci., No. 48983). Herb. Univ. Calif., Berkeley.
Distribution.—Known only from the type
locality.
Merrill's original diagnosis,
translated with additions from his longer English description (enclosed in
brackets), reads as follows: "A shrub 3-4 m high, scarcely aromatic, glabrous,
or nearly glabrous. Leaves [26-] 39 cm long, Leaflets 7-9,
membranaceous, up to 16 cm long and 6.5 cm wide, ovate, elliptic, strongly
inequilateral [at the base], obtusely [acuminate]. Inflorescences
lax, 30-35 cm long, slightly pubescent. Flowers 5-merous, about 5
mm diam. [Calyx shallow, 1.5 mm in diameter, 5-lobed, the lobes
broadly ovate, acute, glabrous, margins sparsely and minutely
puberulent. Petals 5, imbricate, oblong, obtuse, 3.75 to 4 mm long,
2 to 2.5 mm wide, glandular, glabrous. Stamens 10, about 3 mm long;
anthers oblong, apiculate, about 1.5 mm long; filaments somewhat enlarged
below. Ovary globose, glabrous.]"
This species, so far found only on Mindanao Island, P.I., is obviously related
to C. anisum-olens, a species distributed in the Philippines from Luzon
Island to Mindanao Island. However, C. laxiflora, besides
having larger flower clusters, with longer, more slender branches and much
larger leaflets than C. anisum-olens, shows important differences from
the species in the structure of the pistil, as follows: (1) in having the
processes at the tips of the five ovary locules without large oil
glands; (2) in having a much shorter gynophore, which shows a very short basal
annular portion only about 0.2 mm high and 1 to 1.1 mm wide just below the
constriction; (3) in having locular cavities which descend deeply into the
gynophore, reducing its length to 0.6 to 0.8 mm (instead of 1 to 1.2 mm, as in
C. anisum-olens); and (4) in having a large subglobose ovary, nearly
filling the flower bud, which at this stage has almost no distinct
gynophore. Tanaka (1932e, p. 422) reduced this species to a
synonym of C. anisum-olens, but, in view of the striking differences in
the ovary and gynophore characters noted above, it is evident that C.
laxiflora should be retained as a good species.
12. Clausena todayensis Elmer,
Leafl. Phil. Bot. 8:2805. 1915.
Type.—Philippines, Mindanao Island, Davao, Todaya (Mt. Apo) (Elmer, No.
10530). Herb. Bur. Sci.,
Manila.
Distribution.—Known only from
the type, collected on a densely wooded ridge south of the Sibulan River at
1,500 ft. [457 m] altitude.
Elmer's original
description reads, in part, as follows: "Tree, with a 10 m high and 3 dm thick
stem…twigs roughened with lenticels, puberulent when young. Leaves
imparipinnate, thinly coriaceous, paler green beneath, dark green on the upper
conduplicate surface, the acute to acuminate tips strongly recurved…5 to
7-foliolate…leaflets punctate beneath, slightly unsymmetric,
oblongish…obscurely crenate or undulate, the base obtusely rounded, the
terminal leaflet the larger, the basal pair subopposite and much reduced, the
larger ones 7.5 cm long by 3 cm wide at the middle, sparsely pulverulent or
puberulent on both sides; lateral pairs of veins 5 to 9, divaricat…petiolules
only a few mm long, the terminal leaflet upon a 1 to 1.5 cm long rachis
extension. Inflorescence ascending, terminal, paniculate, 10 to 20
cm long; all the stalks terete, green, brown puberulent…the ultimate branchlets
subtended by minute bracts; flowers solitary or usually in small terminal
clusters, subtended by a bract…finely ciliate along the margins; pedicel 1 mm
long, relatively thick; sepals 4, cremeus, caducous, elliptically oblong, 3.5
mm long by one-half as wide across the middle…glabrous…stamens 8, erect;
filaments 2 mm long, fleshy, compressed, purplish-brown sprinkled, pointed at
the apex only…anther 1.5 mm long…emarginate at the apex…ovary more or less
rugose, 2 mm long, short stipitate, obscurely 4-angled and with some large
glands, terminated by a circular stigma."
Tanaka (1932e, p. 423) did not follow Merrill (1923, p. 337) in reducing
this to a synonym of C. anisum-olens because, as he stated, C.
todayensis has a 4-merous "small ovary, with only 8 large oil-glands
somewhat like that of C. harmandiana. The plant is, however,
very gracile and not like the above [C. harmandiana]." Pending further study, the species
is here retained.
13. Clausena
cambodiana (Pierre) Guill. in Lecomte, Not. Syst. 1:219. 1910.
Glycosmis cambodiana Pierre, Flore Forest. Cochinch. 4(18):text to
pl. 285. 1893.
Type.—French Indo-China,
Cambodia, Knang-Krêpeuh Mountains at 1,500 m altitude (Pierre, No.
832). Herb. Mus. Hist. Nat., Paris.
Distribution.—Known only from the type
locality.
The description given by Guillaumin
(1911, p. 665) reads, translated, as-follows: "A small tree 5-8 m
tall. Branches glabrescent. Leaves 15-20 cm long, with
5-11 alternate, glabrous, membranaceous leaflets, equilateral at the base, oval
or oblong-lanceolate, 9-15 X 3-5 cm, cuneiform at the base, cuspidate at the
apex, with 6-9 pairs of lateral veins, which like the veinlets are visible
above and are raised below; petioles cylindrical, pubescent; petiolules
cylindrical, about 5 mm long. Fruiting inflorescence a terminal
panicle, as long as the leaves, pyramidal, pubescent. Fruit
spherical, red-violet, about 8 mm diam., punctate because of the oil glands,
very fragrant."
In the original description
Pierre mentioned that the lower leaflets are smaller and that the fruiting
inflorescences are 20 cm long.
A photograph
of the type specimen (Pierre, No. 832) preserved in the Paris herbarium, made
by Tanaka, shows the leaves taper to a very sharp tip 0.9 to 2 mm wide, and are
very flat and entire-margined. The fruits are borne on slender
pedicels about 4 to 6 mm long, arising from nearly straight, rather stiff
lateral branches of the inflorescence, 7 to 8 cm long below but much shorter
above. Most of the other species of Clausena having caudate
leaflets have the margins of the leaflets minutely dentate or crenulate (as in
C. dentata) or else have curved tips and very unequal-sided bases (as in
C. wallichii).
14.
Clausena brevistyla Oliv. Jour. Linn. Soc. Bot. 5(2):31.
1861.
Type.—Australia, Queensland, Hope
Islets (MacGillivray, July 18, 1848). Herb.
Kew.
Distribution.—Australia:
Queensland.
Oliver's original description
reads: "Leaves 10-15-foliolate, leaflets very oblique; ovary more or less
obovate, narrowed at the base, 4-lobed above, glabrous or almost glabrous;
style distinct from the ovary, very short, half shorter than the ovary, equal
to the stigma [in diam.] Shrub (?). Leaves 5-8 in. [12.5-20 cm]
long. Petiolules about 2 lin. [4 mm] long. Leaflets
oblique or rhomboid-oval or ovate, apex scarcely acuminate, margins
undulate-crenate or dentate, glabrous or in young leaves pubescent near the
midrib, 1-4 1/2 in. [2.5-11.5 cm] long, 10 lin. to 1 1/2 in. [20-38 mm]
wide. Calyx 4-5 parted, lobes widely ovate. Petals 4-5,
thin, broadly elliptical or rounded. Stamens often 10, filaments
thicker below and dilated, somewhat arched. Ovary glabrous, or with
few hairs, 4-5-locular. Ovules 2 (superimposed?). Stigma
equal to the style [in diam.]. Fruit not seen.
Resembles C. heptaphylla very closely, differing in the conspicuous
obliquity of the leaflets and the very short style, inserted in both species in
the central apical depression of the ovary. The flowers are often
pentamerous."
This species is the only one
native to Australia and thus occupies the extreme southeastern portion of the
range of the genus Clausena.
14a. Clausena brevistyla var. halmaheirae
(Miq.) Swing. Jour. Wash. Acad. Sci. 30:82. 1940. Clausena halmaheirae
Miq. Ann. Mus. Bot. Lugd.-Bat. 1:211.
1863.
Type.—East Indian Archipelago;
Moluccas, Halmahera Island (Teijsmann and De Vriese), Rijks Herb.,
Leiden.
Distribution.—Known only from
the original collection.
The original
description by Miquel reads, translated, as follows: "Branches, petioles, and
midribs (on lower side), along with the inflorescences, covered sparingly with
a thin adpressed pubescence; leaves long, with rather widely separated
leaflets; leaflets (on the stronger leaves up to 11, or fewer) with short
petiolules, lower ones nearly opposite, broadly obovate and shortened, the
others not opposite, narrowly sublanceolate-oblong, acutely pointed toward the
base, one side cut off, the other obtusely unequal-sided, chartaceous with even
or slightly repand margins; sometimes very minutely subserrate, pale, black
glandular-punctate below, with 10-15 spreading veins; fruiting inflorescences
axillary, 1/2 longer than the leaves, with lax few-flowered branchlets; calyx
4-lobed; immature berries, 1-seeded. Near to the species which I
listed in Fl. Ind. bat., Suppl. I, p. 501, doubtfully under the name C.
excavata. Sufficiently distinct from all described species, but
recalling several of them. Leaves 1-1.5 ft. [30-45 cm]
long. Lower leaflets 3 in. [7.5 cm] long, upper ones 6-6.5 in.
[15-17 cm] long."
Tanaka (1931b, p. 6)
who examined the type specimen at Leiden and other type material at Utrecht and
Kew, concluded that this plant "is closely related to C. harmandiana,
but is different in having narrower leaflets with distinct black dotting and a
long branched panicle bearing [a] decidedly smaller berry." With
respect to the last-named character, Miquel noted that the type material (all
that is known as yet) has immature fruits.
This variety is characterized by having large fruiting inflorescences with
widely spreading primary branches and sturdy terminal pedicels 2 to 4 mm
long. It has very small pistils, only 1.4 to 1.5 mm long (in dry
state), with a very slender gynophore, only 0.2 to 0.3 mm in diameter, and an
ovary only 0.8 mm long and 0.6 mm wide, rough with oil glands, and a cylindrical
style, shorter than the ovary. The leaflets are only slightly
unequal-sided. This variety is very distinct from C.
harmandiana.
15. Clausena
anisata (Willd.) Hook. f. Niger Fl. 1:256. 1849. Amyris
anisata Willd. Sp. Pl. 2:337. 1800. Illus. De Wildeman, Ann.
Mus. Congo, Bot 5 sér. 2: pls. 52, 53.
1907.
Type.—Western Africa, "Guinea"
(coll. ?). Herb. Bot. Mus.,
Berlin-Dahlem.
Distribution.—Western
Africa: Sierra Leone to Angola; central Africa: Bahr-el-Ghazal and Pemba
through Usambara to northern Malawi and Tanzania on Masai plateau, reaching
3,000 m altitude.
This Species was described
by Oliver (1861, p. 34) from Nigerian material as follows: "Leaflets 10-20,
panicles lax, elongate, shorter than the leaves or longer, lateral branches
divaricate in a cymose manner, slender, pubescent; ovary 4-furrowed, 4-loculed;
ovules paired [in each locule], obliquely superimposed or almost
collateral.…Small shrub 3 ft. [92 cm] high with white flowers.
Leaves 6-12 in. [15-30 cm] long; leaflets unequal-sided, obliquely ovate or
ovate-oblong, obscurely crenulate, sometimes slightly acuminate or emarginate,
pubescent. Inflorescences paniculate, 6-9 in. [15-23 cm] long,
borne in the axils of the upper leaves. Pedicels 2/3-1 lin. [1.2-2
mm] long. Bractioles minute, ovate or lanceolate.
Calyx 4-merous, lobes ovate. Stamens with filaments abruptly
dilated in the middle, subulate above. Ovary short-stipitate,
glabrous. Style rather long, rather thick, 4-furrowed, separating
at the base."
This species is the only one
found in western Africa, but in eastern Africa it shares the central part of
the range of the East African C. abyssinica. All three
African species vary greatly and have ill-defined varieties.
Engler (1931, p. 322) reported that in western
Africa C. anisata is found in the scattered bush forests of the
hinterland of Cameroons (now Cameroon), where it is a tree 15 to 20 m high at
an altitude of 2,100 m. Engler found that in eastern Africa it
reaches the very considerable altitude of 3,000 m (9,840 feet), where it grows
in the mountain meadows as a beautiful tree of medium height with a broad
crown. This is by far the highest altitude yet reported for any
member of the orange subfamily, but since Mount Kilimanjaro is situated only
30° 37' south of the equator, even at high altitudes no severe cold in winter
is experienced there.
Clausena anisata
and C. inaequalis were separated by Engler (1931, p. 322) by the
length of the inflorescences in relation to the length of the leaves: in C.
anisata the flower clusters are much longer than half the length of the
leaves; in C. inaequalis they are never more than half the length of the
leaves. De Wildeman and Durand (1901, p. 743) found the leaves of
C. anisata to be 15 to 40 cm long, those of C. inaequalis to be
7.5 to 25 cm long; and the flowers of C. anisata to occur in
many-flowered paniculate cymes, but those of C. inaequalis in
three-flowered clusters or singly along the racemes. These two
species have similar, more or less obovate or pyriform ovaries, with a large
gland at the tip of each locule.
15a. Clausena anisata var. multijuga Welw. ex
Hiern. Cat. Welw. Afr. Pl. 1:116.
1896.
Type.—Africa, Angola, Golungo
Alto (Welwitsch, No. 1320, in part; coll., Nov. 1855). Herb. Brit.
Mus., London.
Distribution.—Angola:
reported only from Golungo Alta and Pungo Andongo, in elevated stations up to
2,200 ft. [670 m].
Differs from the species
chiefly in having leaves with more numerous leaflets (17-32). A
shrub or "small tree 7-12 ft. [2-3.6 m] high, with slender, far-reaching,
almost sarmentose branches, with a very short, closely-set
pubescence." Leaves 3-10 in. [7.5-25 cm] long; leaflets 1/2-2 X
1/6-5/8 in. [1.2-5 X 0.4-1.5 cm]; inflorescences 4-9 in. [10-22.5 cm] long.
It is doubtful whether this small-leaved
multifoliolate form deserves taxonomic recognition. No mention is
made of tubercules on the ovary such as are found in a form growing a few
hundred kilometers to the north in the Congo Valley; presumably the ovary in
var. multijuga is smooth.
This Angola
form is said to make a small, elegant tree. It should have value as
an ornamental tree for cool subtropical regions. The leaves are
"fragrant when rubbed, emitting a very agreeable aroma like that of oranges and
lemons." The flowers are white but small.
15b. Clausena anisata
var. mollis Engl. Die Pflanzenwelt Ost-Afrikas C:228. 1895; also
in Die. Nat. Pflanzenfam. ed. 2. 19a:322.
1931.
Type.—Africa (locality
doubtful). Herb. Bot. Mus.,
Berlin-Dahlem.
Distribution.—Africa:
forest and bush steppes of Cameroon hinterland; the Lake Region; Tanzania,
Mount Kilimanjaro, western Usambara, coastal region; northern Malawi.
Leaves soft-pubescent and with blunter leaflets
than the species.
An inadequately known plant
that needs study.
16. Clausena
inaequalis (DC.) Benth. in Hook., Niger Fl. 1:257. 1849. Elaphrium
inaequalis DC. Prodr. 1:724. 1824; Amyrisinaequalis Spreng.
1825; Fagarastrum inaequalis Don. 1832; Myaris inaequalis Presl.
1840. Illus. Wood & Evans, Natal Pl. 1(2):54, pl. 66. 1899;
Sim, For. Fl. Cape Col. pl. 26. 1907.
Type.—South Africa (coll. ?). Herb. De Candolle,
Geneva.
Distribution.—Southeastern
Africa: Cape Colony, Pondoland, Natal, with transition forms to C.
abyssinica (fide Engler, 1931, p. 322).
This species was described by Wood and Evans
(1899, p. 54) as follows: "A shrub or small tree with dark coloured bark, which
is usually thickly studded with lenticels. Leaves unequally
pinnate, 6-8 in. [15-20 cm] long; leaflets alternate or subopposite, in 4-6
pairs, petiolulate, glabrous, lateral ones very unequal-sided, the lower
portion of the lamina being much smaller than the upper portion, terminal one
subequal, broadly lanceolate, margin unequally crenate, attenuate to an obtuse
or subacute apex; 1 1/2-2 in. [3.8-5 cm] long; 1/2-3/4 in. [13-18 mm] broad,
petiolules 2-3 lin. [4-6 mm] long, pubescent, terminal one 6-9 lin. [12-18 mm]
long, swollen and bent below the lamina. Flowers paniculate,
white. Calyx small, 4-parted, sepals lanceolate,
pubescent. Petals 4, concave, free, spreading, imbricate in bud,
deciduous. Stamens 8, hypogynous, filaments free, subulate,
flattened. Anthers sagittate. Ovary on a short
cylindrical fleshy torus, obtusely 3-lobed, 3-celled, ovules 2 in each cell,
collateral. Style short, thick, deciduous. Stigma
3-lobed. Berry fleshy, dark purple, the epicarp thickly studded
with minute glands; 1-2 seeded by
abortion."
Clausena inaequalis varies
greatly, as do the other African species, C. anisata and C.
abyssinica. The first two differ in the length of the leaves,
the obliquity of the leaflets, the length of the inflorescence compared to the
length of the leaves, and the number of flowers arising in the ultimate
division of the inflorescence; C. abyssinica is discussed below.
17. Clausena abyssinica Engl.
Die Pflanzenwelt Afrikas 3:757. 1915. Clausena inaequalis var.
abyssinica Engl. Die Pflanzenwelt Ost-Afrikas C:229. 1895.
Illus. fig. 3-4,H.
Type.—Abyssinia, plateau up to 2,300 m altitude (coll. ?). Herb.
Bot. Mus., Berlin-Dahlem.
Distribution.—Eastern Africa: Ethiopia; Tanzania, Mount Kilimanjaro (up to
2,800 m alt.); southern Malawi; Zambia.
The
brief original description of the variety reads, translated, as follows:
"leaflets less unequal-sided, more acuminate [than in the species C.
inaequalis].—3 [Abyssinian plateau] (2,300 m).—15 [Mount Kilimanjaro,
Tanzania] (2,700 m).—A tree up to 10 m high in mountain forests."
The laconic description of the species in Engler's account of Clausena
(1931, p. 322) characterizes C. inaequalis as having "very
unequal-sided, rhombic-ovate, mostly blunt pointed" leaflets, and C.
abyssinica as having "usually oblique-lanceolate leaflets and larger
flowers."
Clausena abyssinica was
reported by Engler (1931, p. 322) as growing up to 2,300 m in the Abyssinian
plateau and up to 2,800 m in the upper girdle-forests of Mount Kilimanjaro in
Tanganyika (now Tanzania). This altitude is exceeded only by C.
anisata, which on Mount Kilimanjaro attains 3,000 m. These two
species hold the altitude record for the orange subfamily, and they must be
able to grow at lower mean and maximum temperatures than any other
Citrus relatives.
A critical
comparative study of C. inaequalis and C. abyssinica is urgently
needed. The original description of C. abyssinica as a
species contradicts the original description of it as a variety in stating that
the leaflets are more oblique than those of C. inaequalis instead
of less oblique! As a matter of fact, all three African species of
Clausena (nos. 15 to 17 above) and their varieties are exceedingly
variable and as yet inadequately described.
Engler (1915, p. 757) noted the difficulty of separating the African species and
varieties of Clausena in an eloquent sentence to the effect that it is as
hard to separate the species as it is easy to recognize the genus, which is
spread over Africa from Ethiopia to Cape Province and from upper Guinea to
Angola.
18. Clausena
papuana Lauterb. Bot. Jahrb. 55:259.
1918.
Type.—Northeastern New Guinea, at
the foot of the Bismarck Mountains (Schlechter, No. 18476). Herb.
Bot. Mus., Berlin-Dahlem; fragments in the Lanterbach Herbarium in the Herb.
Bot. Inst., Univ. Breslau.
Distribution.—Known only from the original collections made by
Schlechter.
Lauterbach's original description
of this species reads, translated, as follows: "Small tree with rounded twigs,
the young shoots tomentose. Leaves alternate, imparipinnate, rachis
tomentose. Leaflets alternate…petiolulate, oblique-ovate, obtusely
subacuminate, the upper half rounded at the base, the lower half acute at the
base, membranaceous, punctate, showing veins on both sides in the dried
material, margins inconspicuously crenulate, lateral veins (8 pairs) oblique,
curving and fusing together near the margins; midrib slightly elevated
below. Panicles axillary, subterminal, as long as the leaves, main
stems tomentose, branches more or less separated, ending in aggregated apical
branchlets, flowers short-pedicellate, in heads (‘capitulate’).
Sepals 4, connate at the base, acutely triangular, tomentose without; petals 4,
elliptic-acute, glandulose, glabrous, reflexed in anthesis; stamens 8, nearly
equal in length, filaments thickened at the base, anthers elliptical,
emarginate; gynophore glandulose; ovary glabrous, cylindric, somewhat
quadrangular, locules 4, each with 2 ovules; style short, stigma
dilated. A small tree with twigs 4 mm diam., having gray-brown
bark. Leaves 30 cm long; rachis 3 mm diam. at lower end; petiolules
4-5 mm long. Leaflets 5-11 cm long, 3-5 cm broad, smallest at the
base of the leaf. Inflorescence 30 cm long, stem 7 cm, branches 2-3
cm, and flower stalks 1-2 mm. Flower buds 2.5 mm diam.; calyx lobes
0.5 mm long, petals 4 X 2 mm. The tip of each petal bears a large
oil gland which, in the bud seen from above, forms a cross with those of the
other petals. Stamens 4 mm long, of which the anther measures 1.5
mm. Gynophore 0.5 mm high, ovary 1 mm high, the style with the
stigma 1 mm long."
Lauterbach noted at the end
of the original description that "this species approaches [C.]
brevistyla Oliv. from Queensland. It differs from it in its
hairiness, larger leaves with broader, blunter leaflets and much larger flower
clusters arising in the axils of the uppermost leaves."
Tanaka (1928c, p. 141) reduced this
species to a variety of C. harmandiana (see p. 177), but, as he pointed
out, this latter species has fewer leaflets (5 to 7 instead of 10 to 11, or
more) and a less hairy rachis. Swingle found the leaflets of C.
harmandiana much less unequal-sided at the base than those of C.
papuana. The flowers of C. harmandiana have remarkably
short pedicels, only about 1 mm long, whereas the pedicels of C. papuana
are usually about 2 mm long. Clausena harmandiana has a very
short, inconspicuous gynophore (Pierre apparently did not see it!), probably
only 1/3 to 1/2 mm long, whereas the gynophore of C. papuana is better
developed, about 1/2 to 3/4 mm long, and is easily seen. Serial
microtome sections made from the type material (Schlechter, No. 18476)
preserved in the Lauterbach Herbarium and kindly lent Swingle by the Botanical
Institute of the University of Breslau show that each locule of the ovary is
tipped by a single large oil gland (not two as in C. harmandiana); each
anther, each petal, and each sepal also has a large oil gland at the tip.
19. Clausena guillauminii Tan.
Bul. Mus. Hist. Nat. Paris, 2 sér. 2:161. 1930. Illus. fig. 3-4,L.
Type.—Thailand, Udawn, Lôti, Wang Sapung (Kerr, No. 8782). Herb.
Kew.
Distribution.—Thailand, Laos.
Tanaka's original description reads, in
translation, as follows: "Shrub; branchlets and leaf veins pellucid-dotted and
puberulous. Leaves 1-5-foliolate, rachis narrowly winged; leaflets
alternate, thick, obovate or oblong, base acute or rather obtuse, apex
cuspidate, obtuse or rather acute, lateral leaflets smaller, nearly
equal-sided, terminal leaflet sometimes much wider, all sparsely black-dotted,
conspicuously reticulate, slightly crenate-serrate. Inflorescences
terminal, paniculate, lateral peduncles short, about equal in
length. Flowers small, 5-merous, petals and calyx lobes
pellucid-spotted at the tips. Filaments subulate, anthers
glandulose-tipped. Ovary ciliate, with 2 series of glandulose
spots, short-stipitate; style long, stigma subcapitate."
The type and cotype material collected in
Thailand by A. F. G. Kerr (his Nos. 8782 and 8388, listed by Tanaka), which was
lent to Swingle by the Director of the Royal Botanic Gardens at Kew, shows the
following dimensions: Compound leaves (No. 8782), 5-7-foliolate, 17-20 X 9-13
cm, rachis 9.5-11 cm long, 1-1.5 mm diam., narrowly margined (0.2-0.5 mm)
between the leaflets; terminal leaflets 7.5-9.5 X 4.3-5.7 cm; lateral veins
9-12 pairs, nearly straight; lateral leaflets 3-8 X 2-4.2 cm.
Unifoliolate leaves (No. 8388), 7.5-10.5 cm long, usually long-elliptical,
blunt or subacute at both ends, petioles 1-3.5 cm long, very narrowly margined;
inflorescences 3-10 cm long, but only 1-2.5 cm wide. This plant has
stems only 20-30 cm high and 3.5 mm diam., springing from a root 1 cm diam.,
yet they were flowering freely and were bearing a few fruits when collected
February 3, 1924.
Craib (1926, p. 233) stated
concerning this material: "The glands on the leaves are by far the largest of
any Siamese rutaceous plant seen. The sepals and petals all have
one prominent gland much larger than the others and the ovary has five
conspicuous glands at the apex and usually a few longish spreading hairs."
Serial microtome sections of a specimen
collected by Kerr (No. 8782) at Me Chan, Chien Sen, Thailand, show that there
are smaller irregularly placed oil glands around the side of the ovary about at
the point where the gynophore ends; there are 7, 8, or more of them, and they,
along with the much larger oil glands that are borne in processes at the tip of
each locule, must be what Tanaka had in mind in describing the ovary as "marked
with two series of glands."
This species is
said to be a shrub growing only 20 to 40 cm tall by Thorel, who collected it in
Laos. Possibly it is killed to the ground by annual fires and
springs up each year from the root. The oil glands are dimorphic,
most of them being small, but a few, scattered 3 to 4 mm apart, being much
larger, l/3 to 1/2 mm in diameter, pale on young leaves but dark brown or
nearly black on old leaves. By transmitted light, such glands are
bright, clear red in color, about Jasper Red of Ridgway (1912, pl.
13). These large oil glands are often conspicuously convex and stand
above the leaf surface; they are sometimes surrounded by many small veinlets
that radiate out from the oil gland.
This
astonishing species merits careful study as it differs widely from almost all
the others of the genus. Two other species, C. papuana and
C. wallichii, have the same distribution of oil glands on the floral
organs, viz., a single, rather large oil gland on the tip of each calyx lobe,
on each petal apex and another [sic] tip, as well as a short-stalked oil
gland at the top of each locule of the ovary. C. papuana has
very different leaves with about twice as many leaflets and a wingless rachis,
whereas C. wallichii has acuminate, crenulate or denticulate leaflets
about twice as large.
20.
Clausena wallichii Oliv. Jour. Linn. Soc. Bot. 5(2):35. 1861.
Illus. Swing. Jour. Wash. Acad. Sci. 30:81, fig. 2.
1940.
Type.—Burma, Chapedong Hill
(Wallich, No. 6370). Herb. Kew.
Distribution.—Burma: eastern Himalayan region, Tenasserim; Thailand.
The original description by Oliver (1861, p. 35)
reads, translated, as follows: "Leaves with the rachis narrowly winged;
leaflets rhomboid-lanceolate, or oblong, acuminate, glabrous, margins
crenulate. Inflorescences paniculate, many-flowered, branches close
together, flowers few. Calyx 5-merous, lobes ovate.
Petals 5, margins slightly imbricate in the bud, thin. Stamens 10,
free, filaments thickened at the middle, at first slightly arched
below. Ovary short and narrowly stalked, 5- (or 4-) locular, ovules
obliquely superimposed. Style very short, furrowed, equaling the
stigma in width." The brief diagnosis adds the following
characters: "Leaves 13-17 foliolate…Style distinct from the ovary, shorter or
almost as long…"
Hooker (1875, vol. 1, p. 505)
added a few characters as follows: "Leaves 8-12 in. [20-30 cm] long; leaflets
2-3 in. [5-7.5 cm] long, alternate, oblique, caudate-acuminate, with an obtuse
notched tip, narrowed into a cuneate base. Panicle 3-5 in.
[7.5-12.5 cm] long, and broad, much branched…Flowers subsessile, 1/8 in. [3 mm]
diam.…ovary glabrous."
An examination of the
type specimen and much other material of this species lent to Swingle from the
Kew Herbarium and from the Calcutta Botanic Garden enabled him to add several
characters, as follows: Leaves 20-30 X 7-12 cm; leaflets 6-10 X 2-4 cm,
crenulate or denticulate, very unequilateral, turning brown on drying, with
numerous evenly distributed oil glands varying greatly in size (about 0.04-0.2
mm diam.); flowers 4.5-5 mm long from base of calyx to tip of stigma, flower
parts all heavily impregnated with tannin residue, glabrous, except ovary;
calyx lobes, petals, anthers, and 5 (or 4) locules of the ovary each tipped with
a single large oil gland; ovary tuberculate, often bearing a few rather long
hairs, about 1.8-2 X 0.8-1.2 mm; style 1.8-1.9 X 0.4-0.5 mm; gynophore 0.7-0.8
mm long, 0.5-0.8 mm diam. at narrowest part; pedicel of flower 1-2 mm long with
minute ciliate bracts at base; fruits subglobose or round-ovoid, 5-6 X 5 mm
(immature?), borne on pedicels 2-3 mm long and 0.8-1 mm thick.
This species has the rachis very narrowly winged
(wings 0.3 to 1 mm wide), a peculiarity it shares with C. guillauminii,
which usually has the rachis merely margined rather than winged, whereas in
C. luxurians the rachis is plainly winged (wings 1 to 2.5 mm
wide). The floral organs, calyx lobes, petals, anthers, and ovule
locules, are each tipped with a single large oil gland, in this character resembling C. guillauminii and C.
papuana. The great variation in the size of the oil glands of
the leaflets resembles but does not equal that found in C.
guillauminii. There can be little doubt that C.
wallichii, C. luxurians, and C. guillauminii are rather
closely related species, and all of them may be distantly related to C.
papuana.
21. Clausena
luxurians (Kurz.) Swing. Jour. Wash. Acad. Sci. 30:79. 1940.
Clausena wallichii var. luxurians Kurz, Jour. Asiat. Soc. Bengal
44(2):133. 1876. Illus. Swing. loc. cit., 30:80,
fig. 1.
Type.—Burma, Pegu, Choungmenah
(Kurtz. No. 1995). Herb. Bot. Gard., Calcutta. A sheet
marked with Swingle and Tillson's serial microtome sections No.
519.
Distribution.—Southern Burma:
known only from Pegu and Tenasserim.
A shrub,
with large leaves, 30-50 X 20-27 cm, 5-7- (often 6-) foliolate; leaflets small
and broad at base of rachis, 6.5-7.5 X 4-5 cm, larger, lanceolate-acuminate
near tip of rachis, 15-21 X 6.5-8.5 cm, base acuminate and decurrent on
petiolules, entire-margined, only slightly unequal-sided, terminal leaflet
symmetrical at base, largest of all, 15-21 X 6.5-9.5 cm, lateral veins varying
greatly in prominence, heavier ones few (7-9), arising at angles of about
70°-75° with the midrib, smaller intermediary lateral veins usually arising at
a much greater angle (often 80°-85°) with the midrib, margins entire or
subcrenate, surface covered with numerous oil glands varying in size from 0.04
to 0.2 mm, rachis narrowly but clearly winged except the segment below the
lowest leaflet, wing broader (1.2-2.5 mm) at top of each rachis segment where
leaflet is attached, smaller (0.8-1 mm) at the base of the rachis segment;
inflorescences usually terminal, short, about 8-12 X 4-6 cm; flowers white,
small, about 4-5 mm diam., 2.5-3.5 mm long from base of ovary to stigma; flower
buds subglobose, all flower parts glabrous, free from tannin, pale
amber-colored when dry; calyx lobes, anthers, and ovary locules each tipped
with a single large oil gland; pistil small, about 2.5-3 mm long; gynophore
well developed, hourglass-shaped, 1 mm long, 0.6-0.8 mm wide at narrowest
point; ovary tubercular at base, 1-1.1 mm long, 1-1.3 mm wide, with one large
oil gland at top of each locule, not blending with the style, which is
narrowest at base, 1.1-1.3 X 0.4-0.6 mm; fruits (immature) subglobose, 5-6 mm
diam.
The abundant material of this plant
collected by S. Kurz at Choungmenah, Pegu (under No. 1995), was labeled by him
C. wallichii var. luxurians. A collection made by A.
Meebold (No. 15090), at Wagôn, Tenasserim (in Herb. Bot. Gard., Calcutta, and
also in Herb. Bot. Gard., Breslau), has somewhat smaller leaves than the Pegu
material and even smaller inflorescences (6 by 3 cm). The fact that
both C. wallichii and C. luxurians have leaves with a winged
rachis, an unusual character in the tribe Clausenae, has probably operated to
cause most taxonomists to class them as mere forms of one and the same
species.
Serial microtome sections of the
pistils show several important differences between these two species in the
absolute and relative size of the flower parts; moreover, C. wallichii
has abundant tannin deposits scattered through the tissues, whereas C.
luxurians shows none. The leaflets of C. luxurians are
decidedly fewer and much larger than those of C. wallichii and differ
also in shape and margination. The flowers of C. luxurians,
however, are decidedly smaller than those of C. wallichii and the petals
are not each clearly tipped by a single large oil gland.
22. Clausena lenis Drake, in
Morot, Jour. Bot. 6:276. 1892. Clausena kerrii Craib, Kew Bul.
Misc. Inform. 1913:67. 1913. Illus. Guillaumin, in Lecomte, Fl.
Gén. Indo-Chine 1:664, fig. 70(6-7).
1911.
Type.—French Indo-China, Tonkin,
Tu-phap (Balansa, No. 3667). Herb. Mus. Hist. Nat.,
Paris.
Distribution.—North Vietnam:
Tonkin; Laos; Thailand: Pâyap; China: Yunnan Province.
A rather full description of this species by
Guillaumin (1911, p. 663) reads, in translation, as follows: "A shrub
completely covered by soft pubescence (except the upper surface of the
leaflets). Leaves 35-40 cm long, with 11-17 alternate leaflets,
very unequal-sided at the base (except the terminal one), membranaceous, oval
(2.5-12 X 2.5-6 cm), acute at the base, obtuse at the tips with the margins
indistinctly denticulate, glabrous above (except the midrib and lateral veins),
completely soft-pubescent below, lateral veins 5-8 pairs, slightly raised on
the underside; veinlets not visible, petiolules about 1 cm long, petiole
cylindric, slightly glandular. Inflorescence a pyramidal,
pubescent, terminal panicle shorter than the leaves, with alternate branches,
pedicels soft-pubescent, as long as the flower, without visible articulations;
flowers 3-4 mm long, flower buds ovoid. Sepals 4, small,
oval-obtuse, slightly pubescent without, ciliate at the margins.
Petals 4, glabrous, oval, glandular. Stamens 8, equal, almost as
long as the petals, anthers lanceolate, with 2 linear locules each ending in a
point, staminal filaments flattened and broadened, very short,
glabrous. Ovary glandular, almost completely glabrous, locules 2,
with 2 superposed ovules in each, style cylindrical, as long as the
ovary. Fruit unknown."
In his
description of C. kerrii, now considered a synonym of C. lenis,
Craib (l.c.) added a few details to Guillaumin's description, as follows:
"Shrub up to 3 m high, twigs up to 5 mm diam., with soft short white
pubescence…Leaves 50 cm long, petiole up to 6.5 cm long, lower leaflets 4.6 cm
long, 3.2 cm wide, upper leaflets up to 14 cm long and 6 cm
wide…Inflorescences…bracts about 1.5 mm long…Petals white (fide Kerr),
oblong-ovate, 5 mm long, 2.7 mm wide. Stamen filaments 0.75 mm
long, anther 3 mm long. Ovary 1.25 mm high, style 2.5 mm long,
provided with a very few hairs, here and there."
The Chinese form may be described as follows:
Small tree, 2-3 m high; twigs stout, 5-6 mm diam., at first angled, soon
becoming terete, finely puberulent, soon glabrescent; leaves large, 30-65 cm
long, with 11-15 leaflets (usually 13-15), rachis minutely puberulent; leaflets
very unequal-sided at base, oblong-lanceolate and acuminate or even subcaudate,
cuneate at base, varying greatly in size on the same leaf, lower ones sometimes
only 2-2.5 X 1.5-2 cm, those in the middle of the leaf largest, 10-13 X 3.5-5
cm, terminal leaflets 8-10 X 2-3.5 cm, lateral veins 6-8 on apical half and 5-6
on basal half on the larger leaflets, margins dentate, except near the base,
midrib and lateral veins minutely puberulent, petiolules 2-4 X 1 mm, leaf
margins more or less decurrent on sides; inflorescences terminal, or axillary
among reduced leaves at the end of the flowering branch, 15-28 cm long and
10-15 cm wide when terminal, 5-15 X 2-4 cm when axillary, ultimate branchlets
bearing pedicels 4-8 X 0.5 mm in dry state, entire inflorescence minutely
puberulous; flower buds ovoid, light coffee brown in dry state; calyx very
small, 4-lobed, lobes semicircular, 0.8-0.9 mm wide, 0.5-0.6 mm long,
puberulous; corolla 4-merous, petals thick, coriaceous, entire, incurved at
tips, 4-5 X 3-4 mm, glabrous, dotted with small oil glands; pistil 4 mm long in
the flower bud, 5-6 mm long after petals and stamens fall; gynophore 1 mm high
with a short annulus at the base 1.2 mm wide, 0.35-0.5 mm high, narrow
constriction above annulus 0.8-0.9 mm wide, upper part of gynophore
cylindrical, 1.2 mm wide, 0.7-0.8 mm high, merging with the equally wide base
of ovary at top but with a complete ring of 12-15 oil glands at the base just
above the constriction; ovary cylindrical below, 1.2 mm diam., slightly narrowed
and rounded at top where a pit, 0.5 mm wide and 0.2-0.3 mm high, bears the
style, slightly narrowed where attached, surface covered with numerous slightly
prominent oil glands and sparingly hirsute, style cylindrical, 0.5-6 mm diam.,
and 3.5-3.8 mm long with no clearly marked stigma, ovules collateral, 2 in each
locule. Staminal filaments very short, anthers very long (4-5 mm),
slender with a single elongated oil gland at back where filament is attached;
fruit (immature) subglobose or slightly ovoid, strongly dotted with oil glands,
5-8 mm diam.
In Thailand, this species has
scattered hairs toward the base of the style. The ovary is said to
be entirely glabrous, but in Tonkin, North Vietnam, it is apparently
occasionally slightly hairy, to judge from Drake's description. In
Yunnan Province, the ovary usually bears scattered, rather long hairs which
often fall away as the young fruit develops, but the style is usually
completely glabrous. The type specimen from Tonkin shows rather
blunt-pointed leaflets with undulate margins; the Siamese material (C.
kerrii Craib) has acutely pointed or acuminate leaflets with faintly and
distantly serrulate margins; and the Chinese plant shows acuminate or caudate
leaflets with rather strongly dentate-serrulate margins. The length
of the style varies somewhat also, being shortest in the Tonkin type and
longest in the Chinese material.
This
remarkable species is very different from any others of the genus, as noted above; it has evidently gone some distance on an independent line
of evolution distinct from that followed by any other known member of the tribe
Clauseneae.
23. Clausena
suffruticosa (Roxb.) Wt. & Arn. Prodr. 96. 1834. Amyrissuffruticosa
Roxb. Fl. Ind. 2:250. 1832.
Type.—None found; type plant sent from Chittagong, northwestern Burma, to
Calcutta Bot. Garden.
Distribution.—India: eastern Himalayan region, Khasi Hills; Burma:
Chittagong.
An undershrub with simple stems,
often only 1-2 ft. [30-60 cm] tall, with all the younger parts softly
pubescent; leaves 30-40 cm long; leaflets 11-17, densely soft-pubescent when
young, when older glabrescent, lower ones ovate, equal-sided, subopposite, 2-4
X 1-2 cm, with entire margins, upper ones alternate, slightly unequal-sided,
ovate-oblong to rhomboid-elliptical, more or less accuminate, sometimes acute,
sometimes blunt at the tip; inflorescences axillary simple or slender panicles,
shorter than the leaves; flowers 4-merous, small, greenish-white, borne on
rather long (3-4 mm), slender, hairy pedicels; petals about 4 mm long; pistil
3-4 mm long; ovary ovoid, 1.5-2 mm long, 1.1-1.6 wide, not borne on a slender
stipe, 4-locular with small oil glands but no large one at the top of each
locule; style short, cylindrical, 2 mm long, 0.4-0.8 mm wide, tip truncate,
stigma not marked; fruits oblong or fusiform, nearly 2.5 cm long by 0.8-1 cm
wide, drooping, glandular, orange-colored, succulent, 1
seeded.
C. suffruticosa is a remarkable
undershrub that needs further study. It is evidently deciduous in
the Chittagong Hills in northwestern Burma, as the young leaves and flowers
appear together in early spring. The twigs are often stout, 7 to 8
mm in diameter, with pith 4 to 5 mm in diameter; the leaf scars of the previous
year are heart-shaped, 6 to 7 mm long and 5 to 6 mm broad.
Apparently the simple short stems arise from creeping rootstocks, perhaps
because of annual fires killing the older wood to the ground.
The pistil of this plant when studied in detail
by means of serial microtome sections made by the modified Juel method (see
Swingle, 1939b, p. 270) shows that it differs strikingly from most of the
other species of Clausena in having no well-developed gynophore on which
the ovary is borne and no cells filled with tannin residues scattered through
the tissues. In this latter character, C. suffruticosa
resembles C. luxurians, which does, however, have a definite, though
small, gynophore. It also differs in several other important
characters.
Clausena dentata var.
longipes has a pistil resembling that of C. suffruticosa in
general morphology; i.e., it has a poorly developed gynophore and a rather long
style merging gradually with the apex of the ovary. However, cells
containing tannin residues are found sparingly in the peripheral tissues of the
ovary of C. dentata var. longipes and abundantly in the
integuments of the ovules.
The fruits of C.
suffruticosa, which were described by Kurz (1877, vol. 1, p. 190) as
"oblong, drooping, nearly an inch long, one-seeded, orange-coloured,
succulent," are very different from those of any other species of
Clausena.
IV. Murraya Koenig
IV. Murraya ("Murraea") Koenig ex Linn. Mant. Pl.
2:554. 1771; nomen conservandum, Regles Intern. ed. 2. 91. 1912.
Chalcas L. 1767; Bergera Koen. 1771; Murraya Koen. 1774;
Marsana Sonner. 1782.
Type
species.—Murraya exotica L. = M.
paniculata.
Distribution.—India;
Indo-China; Thailand; southern China: Hainan; Taiwan; Malay Peninsula; Andaman
Islands; Sumatra; Java; Borneo; Philippines; Celebes; New Guinea; New
Caledonia; northeastern Australia.
Common
name.—Orange jessamine (or murraya).
Unarmed trees with odd-pinnate leaves with alternate leaflets; inflorescences,
rather large panicles, either axillary or terminal; flower buds cylindrical or
long-ovoid; flowers rather large, 5-merous; calyx of 5 ovate or lanceolate
sepals united at the base or only in the lower third; petals 5, rather large,
lanceolate or linear, imbricate; stamens 10, free, elongate, filaments
flattened in some species, anthers small, broadly elliptic or oval; disk
annular, cushion-shaped or cylindrical, short; ovary ovoid, with 2-5 locules,
each with 2 (or 1) superimposed or almost collateral ovules; style rather long
and slender, finally falling off, stigma capitate; fruit a small berry, ovoid
or subglobose, with mucilaginous pulp; seeds medium-sized, with a thin testa,
cotyledons plano-convex.
Murraya is a
genus containing eleven species, of which all but three are rather closely
related species that fall into two or three groups not as yet adequately
studied. The only relationship of Murraya that appears clear
is with Clausena, a genus that is difficult to distinguish at first sight
from Murraya except by its having, as a rule, much shorter styles and
smaller flowers with larger anthers. Careful study of the genus
Clausena has, however, shown that all the species have an
hourglass-shaped gynophore of a character not found in
Murraya. The tissues of all the flower parts in
Clausena are usually highly impregnated with tannin residues deposited in
some but not all of the cells. The curious species M.
alternans, which seems to be related to M. alata, has a stalked ovary
unique in Murraya but of different morphology from the gynophore found in
Clausena.
The anomalous species M.
stenocarpa, having unifoliolate leaves, is still too inadequately known to
permit discussion of its relationships.
It is
clear that Murraya is not closely related to Citrus; nevertheless
Citrus has at least once been successfully grafted on M.
paniculata, a vigorous species easily propagated from cuttings.
Dr. Toxopeus, working in Java (1936, p. 6), reported having crossed
Citrus and M. paniculata, but he obtained only stunted
hybrids.
Tanaka (1929a) published a
monographic study of the species of Murraya (under the genus name
Chalcas) which has greatly helped in understanding this
genus. He was the first taxonomist to transfer Micromelum
glabrum Guill. and Atalantia stenocarpa Drake to the genus
Murraya.
A key to the eleven species of Murraya is presented.
1. Murraya paniculata (L.)
Jack, Malay. Misc. 1:31. 1820. Chalcas paniculata L. Mant. Pl. 68.
1767; C. camuneng Burm. f. 1768; Murraea exotica L. 1771;
Murraya exotica L. 1774; Marsana buxifolia Sonner. 1782;
Limonia lucida Forst. 1786; Murraya sumatrana Roxb. 1832; M.
odorata Blanco, 1845; M. scandens Hassk. 1866; Chalcas exotica
(L.) Millsp. 1895; Murraya banati Elmer, 1915. Illus. Murray,
Comment. Soc. Reg. Göttingensis 9:186-91, pl. 1. 1789; Bellenden-Ker,
Bot. Reg. 5:pl. 434 (col.). 1819; Hooker, Exot. Fl. 2:pl. 134.
1825; Wight, Icon. Pl. Ind. Or. 1:pl. 96. 1840; Beddome, Fl. Sylv. Anal.
Gen. pl. 7, fig. 2. 1871; Penzig, Studi Bot. Sugli Agrumi, Atl. pl. 17, fig.
15; pl. 18, figs. 12-16. 1887; Guillaumin, in Lecomte, Fl. Gén. Indo-Chine
1:665, fig. 69(5). 1911; and many others (see Stapf, Index
Londinensis 4:333 [1930]); fig. 3-7 this work.
Type.—British India. Linn. Herb. London (substitute type not
labeled by Linnaeus, fide Jackson, 1912, p,
106).
Distribution.—India, Ceylon,
Burma, Indo-China, China, Taiwan, Malay Peninsula, East Indian Archipelago,
Philippines, Australia, New Caledonia, Melanesian
islands.
Common name.—Orange
jessamine.
Kurz (1877, vol. 1, p. 190)
described this species as follows: "An evergreen tree, 15-25 ft. [4.6-7.6 m]
high, trunk 6-8 ft. [1.8-2.4 m] high, 1 1/2-2 ft. [46-61 cm] diam., the young
shoots puberulous; leaves unpaired-pinnate or occasionally pinnately
3-foliolate, glossy, glabrous, or sometimes the rachis puberulous; leaflets
alternate, cuneate-obovate or almost obliquely rhomboid, shortly petioluled,
blunt or bluntish acuminate, 1-1 l/2 in. [2.5-3.8 cm] long, coriaceous; flowers
rather large, white, in dense but small, almost sessile terminal corymbs;
petals about 1/2-3/4 in. [13-18 mm] long, recurved; stamens 10, alternately
shorter; ovary 2-celled, the style long with a capitate glandular stigma;
berries ovoid-oblong, bluntish acuminate, nearly 1/2 in. [13 mm] long,
orange-coloured, 1-2-seeded; seeds villous."
This species is very widely distributed and doubtless extremely
old. However, it has evolved at a very slow rate, as is shown by
the small morphologic differences that distinguish it from var.
ovatifoliolata (noted below), which has been separated from the other
forms of the species for many millions of years ever since Australia lost all land connections in the Eocene period. The
wood of this species—particularly that of the roots—is much valued by the Malays
for making kris handles, Burkill (1935, vol. 2, p. 1507) noted that the trunks
split unless carefully seasoned and that few pieces are larger than 8 inches
(20 cm) in diameter.
The orange jessamine is
a handsome greenhouse ornamental that blooms profusely. It has
large white fragrant flowers that are succeeded by small red fruits (see fig.
3-7). In warm subtropical climates it thrives out of
doors. It grows vigorously and can be propagated easily from
cuttings. Citrus cannot as a rule be grafted on it, although
one strain of lemon (an unidentified seedling) made a good growth on it in the
greenhouses of the former Bureau of Plant Industry at Washington,
D.C. There are many varieties and strains of M. paniculata
in the Old World; possibly some of them would support Citrus better
than the strain now grown in the United States, probably originally introduced
from India.
1a. Murraya
paniculata var. ovatifoliolata Engl. Die Nat. Pflanzenfam.
3(4):188. 1896. Murraya ovatifoliolata (Engl.) Domin, Bibl. Bot.
89:296. 1927. Illus. Bailey, Compr. Cat. Queensl. Pl. fig. 61
bis. 1909.
Type.—Queensland (F.
M. Bailey). Herb. Bot. Gard., Brisbane
(?).
Distribution.—Australia:
Queensland.
Engler distinguished this variety
from the species as having broadly oval or ovate leaflets. Bailey
(l.c.) described this form as follows: "This, our indigenous form, is of a more
straggling habit with more numerous and larger oil-dots, and is often decidedly
hirsute and tomentose, thus very distinct from the two Indian ones of our
gardens." The leaves are 3-9-foliolate; the twigs, calyx, petals,
and ovary hirsute.
The essential oil in the
leaves of var. ovatifoliolata from Queensland was compared with that from
the species growing at Dehra Dunn, India, by Penfold and Simonsen (1925, pp.
146-55). The oil of both species and the variety was found to
consist chiefly of sesquiterpenes and sesquiterpene alcohols, but "the chief
constituent of the Indian oil was found to be l-cadinene…whilst
bisaboline was similarly identified as one of the sesquiterpenes present in the
Australian oil." The other constituents were not identified as
they did not yield any crystallizable derivatives. Significant
differences were also found in the saponification value of the two
oils. The leaves of the species grown in India yielded on steam
distillation 0.01 per cent of dark-colored oil with a saponification value of
8.87; leaves of the variety from Queensland yielded 0.06 to 0.15 per cent of
oil having saponification values of from 19.37 to 26.18. There were
also differences in the rotative power of the oils, that of the species being
[a]D-78.2°; that of the variety from -10.2° to -12.2°.
1b. Murraya
paniculata var. zollingeri Tan. Jour. Ind. Bot. Soc. 16:232.
1937. Chalcas paniculata var. zollingeri Tan. Bul. Soc. Bot.
France 75:709. 1928.
Type.—Bima
[or Sumbawa] Island (Zollinger, No. 3351). Herb. Mus. Hist. Nat.,
Paris.
Distribution.—Central East
Indian Archipelago; Bima, Sumbawa, and Timor islands.
Leaflets 3-6, small, chartaceous, margins much
reflexed; rachis thin, declinous, slightly pubescent; fruits not apiculate,
often pendulous, sparsely puberulous.
Tanaka
(1929a, p. 27) is uncertain whether this is not the same as M.
heptaphylla Spanoghe, in Linnaea 15:178 (1841).
1c. Murraya paniculata
var. omphalocarpa (Hay.) Tan. Jour. Soc. Trop. Agr. 1:27. 1929.
Murraya omphalocarpa Hay. Icon. Pl. Formos. 3:51.
1913.
Type.—Formosa, Totosho (coll.,
Kawamakami). Herb. Imp. Univ.,
Tokyo.
Distribution.—Known only from
Taiwan.
Differs from the species in the
following characters: fruits large with attenuate tips, about 21 X 12 mm (when
immature, rostrate, about 15 X 5 mm); flowers larger; petals narrowed at the
base; calyx lobes elongate, ovate to linear-oblong, 3 mm long; leaflets broad,
5-8 X 4-4.7 cm.
Tanaka (1929a, p. 28)
stated that "these characters may be linked by gradations" with those of the
species, but as the differences are fairly large they can safely be regarded
as varietal but not specific.
2.
Murraya gleniei Thwaites ex Oliv.
Jour. Linn. Soc. Bot. 5(2):29. 1861. Chalcas gleniei (Thwaites)
Tan. Bul. Soc. Bot. France 75:710. 1928. Illus. Trimen, Handbook
Fl. Ceylon, Atl. pl. 22. 1893.
Type.—Ceylon (Thwaites, No. 3627). Herb. Kew.
Distribution.—Known only from
Ceylon.
This species was best understood by
Trimen (1893, vol. 1, p. 220), who, while he was director of the Botanic
Gardens of Ceylon (where alone the species is found), naturally had abundant
material at his disposal. His description reads: "A bush, much
branched, bark very white, young twigs strongly pubescent; leaves
imparipinnate, 1 1/2-4 in. [3.8-10 cm long]; rachis strongly pubescent,
leaflets 3-7, on short pubescent stalks, upper ones much the longest,
rhomboid-lanceolate, obtuse, emarginate, lower ones rotundate, all slightly
crenate, glabrous; flowers 1/2-3/4 in [13-19 mm] diam., about 3-7, in short,
racemose, axillary and terminal cymes, pedicels very pubescent; sepals very
small, densely tomentose; petals 1/2 in. [13 mm long]; ovary on a distinct
cylindrical stalk, 5-lobed, 5-celled; berry large, 1 in. [diam.], somewhat
pyriform or urn-shaped, flat-topped, but mammillate in centre, 5-lobed or
bluntly 5-angled, rough with large glands, shining green, 5-celled, the large
inflated cells each containing 1-3 globose green seeds."
This Ceylonese form of the orange jessamine is
much like the normal form of the species except in its curious inflated
star-shaped fruits. It may perhaps be a mutation which has been
able to establish itself as a satellite species. Engler (1931, p.
320) noted that the ovary is "4- or 5-locular" and consequently the fruits must
be also 4- or 5-lobed.
3.
Murraya alternans (Kurz) Swing. Jour. Wash. Acad. Sci. 28:532.
1938. Limonia (?) alternans Wall., in Voigt, Hort. Suburb. Calcut.
139, nomen nudum. 1845; L. alternifolia ("Wall. ap. Voigt") Kurz,
Jour. Asiat. Soc. Bengal 42:64. 1873; L. alternans ("Wall.") Hook.
f. Fl. Brit. Ind. 1:508, 1875; L. alternans ("Wall.") Kurz, For.
Fl. Brit. Burma 192. 1877.
Type.—Burma,
Pegu (Kurz, No. 2010). Herb. Kew.
Distribution.—Known only from type
locality. Kurz (1877, vol. 1, p. 192) stated: "Not unfrequent in
the upper mixed and occasionally in the moist forests of the Pegu Yomah
[Mountains, about Lat. 18°40'-19° N., Long. 96°35'-97° E.], sporadical but
usually gregarious."
A slender, simple or
sparingly branched shrub, 60-120 cm [2-4 ft.] tall, with deciduous foliage;
twigs slender, glabrous, with numerous shallow, longitudinal furrows and low
ridges; leaves odd-pinnate, completely glabrous, 11-15 foliolate; leaflets
alternate, nearly sessile, oblong-lanceolate to lanceolate, 2.5-4 cm long,
somewhat unequal-sided toward the cuneately narrowed base, slightly attenuate
to a blunt tip, margins slightly crenate above the middle; rachis narrowly
winged; inflorescences small, short-peduncled, glabrous, axillary cymes,
appearing with the new leaves; flower buds small, oblong or elliptical, 3-5 X
2-3 mm, flowers short-pedicellate; calyx nearly flat, 5-lobed, lobes
triangular, glabrous, 1-1.5 mm long, 1.5-2 mm wide, with numerous oil glands;
petals 5, white, upright, linear, 5-7 mm long, tapering to a point, with many
oil glands in the upper half; stamens free, filaments 10, slightly swollen
above, sparingly short-pilose, anthers small, oblong, 1.2-1.3 mm long; pistil
glabrous, 4.5-5.5 mm long; ovary borne on a long, slender, nearly cylindrical
stipe, 0.8-1 mm long and 0.4-0.5 mm wide, arising from flat top of the disk
(said to elongate after flowering), the ovary itself obovate, 1.8-2 mm high,
1.1-1.2 mm wide, bluntly rounded or obcordate at top because of 2 large oil
glands, 1 over each locule; locules 2, each with 2 ovules suspended from the
top; style slender, subclavate, 2.2-2.6 mm long, 0.2-0.5 mm wide, stigma at tip
not clearly delimited; disk annular, as broad or broader than the ovary, 1.8-2
mm wide, 0.5-0.6 mm high, fruits ovoid, glabrous, short-stalked, about 4 mm
long.
This species was assigned, somewhat
doubtfully, to Limonia by Kurz and Sir Jose Hooker, but has apparently
never been studied since the 1870's. It is no longer mentioned in
lists of plants covering Burma, and even the genus Limonia, to which it
was assigned when discovered (and from which it has never been removed), is no
longer in use (see Swingle, 1914-17, pp. 327-28).
This extraordinary plant, frequently producing
only a single slender stem, 2 to 4 feet high, is unique in the genus
Murraya in having deciduous foliage. This species together
with Clausena pentaphylla, C. dentata var. dulcis, and
C. suffruticosa are the only members of the tribe Clauseneae having
deciduous foliage. The leaves drop off during the hot, dry season,
which in Pegu begins toward the end of February and lasts until
May. When the summer rains come on, new leaves push out on the
bare stems, and with them clusters of small white flowers appear in the axils
of the leaves.
The type specimen of this
species, collected in Pegu in 1872 by Kurz, was kindly lent to Swingle by Sir
Arthur Hill, director of the Royal Gardens at Kew. A study of the
flower anatomy from serial microtome sections made from this Herbarium material
by the modified Juel method shows clearly that it belongs in the genus
Murraya and is a species near to M. alata, which occurs not far to
the eastward in Indo-China and which also has leaves with a winged
rachis. It differs from all the other specie of Murraya in
having the ovary borne on a slender, nearly cylindrical stipe.
Kurz, in publishing his first description of this
species in 1873, mistakenly used the specific name "alternifolia,"
although he professed to recognize the specific name "alternans" first
printed (but not adequately published) by Wallich in Voigt in 1845.
Hooker corrected this misspelling in 1875 and Kurz accepted the correction in
1877.
4. Murraya alata
Drake, in Morot, Jour. Bot. 6:276. 1892. Chalcas alata (Drake)
Tan. Jour. Soc. Trop. Agr. 1:35.
1929.
Type.—French Indo-China, Tonkin,
Tan-keuin (Balansa, Nos. 1118, 1119).
Distribution.—North Vietnam: Tonkin (Tan-keuin, Kien-khé, and Bac-bat),
Annam.
The brief original description reads,
in translation, as follows: "Shrub, 1 m high. Leaves up to 10 cm
long, completely glabrous; petiole and rachis alate; leaflets 7, 3-4 cm long,
1-2 cm wide, oblong or rhomboid, acute or subacute at the tip, obscurely
crenate; petiolule very short and inconspicuously puberulous. Cymes
axillary, 2-3 cm long, few-flowered, puberulous. Sepals
acute. Petals oblong. Staminate filaments
linear. Berry ovoid."
This
description was based on the type material from Tan-keuin, Tonkin, North
Vietnam (Balansa, Nos. 1118, 1119). Guillaumin (1911, p. 659) added
a few characters not mentioned by Drake del Castillo, since he had additional
material at his disposal from Kien-khé and Bac-bat, Tonkin, collected by
Bon. The more important characters first noted by Guillaumin are
described, in translation, as follows: "…Branches glabrous, with yellow
bark. Leaflets 5-7…coriaceous, entire or undulate on the margins,
lateral veins scarcely visible. Inflorescences…2-3 cm long,
branches 15 mm long, pedicels 9 mm long. Sepals…triangular, acute,
1.5 mm long, pubescent without. Petals glabrous, erect, obovate,
10-15 mm long, with parallel veins. Stamens 10, the 5 longest a
little shorter than the petals, and reaching to the stigma; filaments glabrous,
subulate; anthers very small, attached at the base…terminated by a small grand,
not easily visible. Disk annular. Ovary glabrous,
ovoid; style subulate; stigma capitate; locules 2 each with a single
ovule. Fruits red, grouped in corymbs, fleshy, with 1 or 2 seeds
which have a parchment-like testa and a single-straight embryo with fleshy
plano-convex cotyledons."
Like M.
alternans, which it resembles in having leaves with a narrowly winged
rachis, this species is reported as being a shrub only 1 m high.
M. alata differs from M. alternans in having persistent instead of
deciduous leaves, and flowers with petals about twice as long. The
calyx lobes of M. alata are pubescent without and the staminal filaments
are glabrous, both conditions being just the opposite of what is found in M.
alternans.
4a.
Murraya alata var. hainanensis Swing. Jour. Wash. Acad. Sci.
32:26. 1942.
Type.—China, Hainan
Island, strand, Haichow (McClure, No. 7611). Herb. Natl. Arbor.,
Washington, D.C.
Distribution.—China:
known only from Hainan Island.
This variety
differs from the species found in Tonkin and Annam in North Vietnam in having
the leaflets when young minutely puberulent on both surfaces and on the
margins. As they mature, the leaflets often become nearly
glabrous, but the petiolules and the rachis remain more or less pubescent even
on mature leaves, and often scattered, short cilia may be still seen along the
margins near the base of mature leaflets. The calyx lobes are
pubescent and the staminal filaments are glabrous as in the
species. The leaflets seem to be rather more variable in shape
(especially in width) than those of the Indo-Chinese typical form.
The fruits are ovoid, about 9-11 X 6-8 mm in herbarium specimens.
This variety is a shrub growing from 1 to 2.5 m
high along the beaches, and in the forests of Hainan from sea level to 1,300 m
altitude. The fruits are said by one collector to be "lustrous deep
red" when ripe.
5. Murraya
koenigii (L.) Spreng. Syst. Veg. 2:315. 1825. Bergera koenigii
L. Mant. Pl. 563. 1771; Murraya foetidissima Teijsm. & Binn.
1864; Chalcas koenigii (L.) Kurz, Jour. Asiat. Soc. Bengal
44(2):132. 1875. Illus. Wight, Icon. Pl. Ind. Or. 1:pl.
13/334. 1838; Guillaumin, in Lecomte, Fl. Gén. Indo-Chine 1:655, fig.
69(4). 1911; figs. 3-8 and 3-9 this work.
Type.—Eastern British India (Koenig, suppl. type). Linn. Herb.,
London.
Distribution.—Indian peninsula;
northeastern and northwestern India; Ceylon; Burma; North Vietnam; South
Vietnam; Cambodia; Laos; China: Hainan Island, Yunnan, and
Kweichow.
Common name.—Curry leaf.
Kurz (1877, vol. 1, p. 190) described this
species as follows: "An evergreen tree, 15-20 ft. [4.6-6.1 m] high, trunk 4-10
ft. [1.2-3 m] high, 1/2-1 1/2 ft. [15-46 cm] in circum., glabrous or slightly
puberulous; leaves unpaired-pinnate, the rachis usually more or less pubescent,
rarely quite glabrous; leaflets in 5-10 pairs with an odd one, on a short
puberulous petiolule, oblong-lanceolate or ovate, almost falcate, oblique at
base, about 1-1 1/2 in. [2.5-3.8 cm] long, acuminate, more or less serrulate,
membranous, glabrous, except on the midrib, which is often puberulous; flowers
small, white, in terminal corymbs; petals oblong-lanceolate, acute, about 2-3
lin. [4-6 mm] long; stamens 10, alternately shorter; ovary 2-celled, the style
short and thick; berries oblong, somewhat acute, the size of a small pea,
l-2-seeded, bluish-black."
This species was
said by Trimen (1893, vol. 1, p. 219) to be "extremely like the small-leaved
form of Micromelum pubescens [= M. ceylanicum], from which it may be distinguished by its more
numerous more pubescent and small leaflets, and more compact corymbose terminal
inflorescence." The pistil is much shorter than the stamens.
Dr. David Fairchild reported (Bur. Pl. Ind.,
Inventory 88, No. 68351) that "the fresh leaves form a constant ingredient of
the Ceylonese curries and give them a very agreeable flavor." He
stated further (1930, p. 284) that the fresh leaves of this species are "deemed
essential to all the curries of Ceylon." They are "boiled with the
curry but thrown out of it before serving." Dr. Fairchild had a
fine fruiting tree of M. koenigii growing in his tropical garden, "The
Kampong," at Coconut Grove, Florida, where it was one of the most prized of his
collection of citrus fruits and their wild relatives (see fig. 3-8). He regularly used the fresh leaves in
making curry.
Trimen stated (1893, vol. 1, p.
221): "This is the familiar 'curry-leaf,' a constant ingredient in curries and
mulligatawny." He reported the species as rather rare in the wild
state in Ceylon but "very much cultivated," doubtless for use in curries.
Dr. David Fairchild obtained seeds of this
species in 1926 in Ceylon. Plants grown from this seed have thrived
on the peculiar soil (a very sandy loam intermixed with porous limestone
fragments of rock that often occupy half or more of the bulk of the soil) found
in Miami and vicinity in the extreme southeastern part of Florida.
6. Murraya microphylla (Merr.
& Chun) Swing. Jour. Wash. Acad. Sci. 32:26. 1942. Clausena
microphylla Merr. & Chun, Sunyatsenia 2:251. 1935. Illus.
Merrill & Chun, loc. cit. fig. 27.
1935.
Type.—China, Hainan Island,
Sunchuen (Chun and Tso, No. 43323). Herb. Arnold Arbor., Harv.
Univ., Cambridge.
Distribution.—China:
known only from Hainan Island.
Common
name.—Small-leaved murraya.
The original
description by Merrill and Chun reads, condensed and translated, as follows: "A
shrub or small tree, branches glabrous, branchlets with a dispersed white
pubescence;…leaves imparipinnate…5-7 cm long, petiole and rachis with
scattered white pubescence of short curved hairs; leaflets, 5-10 on each side,
alternate or subopposite…oblong or oblong-ovate or oblong-lanceolate, upper
ones usually larger up to 2 cm long…lower ones subopposite, 3-4 mm diam.;
intermediate ones 5-18 mm long, 3-6 mm wide, all of them crenulate, obtuse…base
usually inequilateral; lateral veins 3-6 pairs…inflorescences terminal,
sparingly pubescent…short-pedunculate, 2-2.5 cm long and wide; [flowers
unknown]; fruits few, ellipsoid or ovoid-ellipsoid, up to 1 cm long, glabrous,
glandular-punctate…persistent sepals 5, oblong-ovate, punctate, glabrous,
obtuse or subacute, 1 mm long; pedicels scatteringly pubescent, 2-2.5 mm
long."
A specimen collected by C. Wang (No.
32740), July 4, 1933, on the Fourth Hainan Expedition of Sun Yatsen University
(in Herb. Arnold Arbor.) has good flower buds, 4-5 X 2.5 mm: petals show
scattered, very dark oil glands. Serial microtome sections show the
pistil to be 4 mm long, disk very short and not delimited from the ovary, which
is 1.2-1.5 mm long and 0.9-1 mm wide, with 1 or 2 locules, with 1 or 2 large
oil glands at the top of the locule (not in a locular process), merging into
the broad style, about 3 X 0.4-0.5 mm, slightly swollen at the top into a
subcapitate stigma, 0.7-0.9 mm diam., which shows a few small oil
glands. The tips of the petals also show several fairly large oil
glands. The stamens are about as long as the pistil, with linear
subulate filaments.
The finding of flowers of
this interesting plant shows clearly that it is a species of the genus
Murraya, of which it is a striking member, very different from any other
species because of its small leaves and tiny leaflets. The
collector's notes on the Hainan specimens show that it may reach 6 m in
height, and have a trunk diameter of 15 cm. It bears white
fragrant flowers.
7. Murraya
siamensis Craib, Kew Bul. Misc. Inform. 1926(8):340. 1926. Chalcas
siamensis (Craib) Tan. Bul. Soc. Bot. France 75:710.
1928.
Type.—Thailand, Lampang, Mê Kat,
alt. 250 m (Winit, No. 849). Herb.
Kew.
Distribution.—Thailand. Reported from four administrative
districts by Craib (1926, vol. 1, p. 230).
Craib’s original description, translated, reads as follows: "Small tree, trunk
with fuscous bark having high ridges (fide Winit); young twigs with
curly pubescence, year-old twigs puberulous, then fuscous, with small
lenticels. Leaves 15-24 cm long (including the petiole, 1-2.5 cm
long), petiole and rachis almost terete with dense curly pubescence; leaflets
17-25, alternate or rarely subopposite, inequilateral (one half oblanceolate or
lanceolate, narrowed at the base; the other half ovate, subtruncate at the
base), apex obtuse, subacuminate, 2-7 cm long, 1-2.7 cm broad, chartaceous,
above curly pubescent, very soon more or less glabrescent below, chiefly
pubescent at the midrib, lateral veins 5-7 pairs, anastomosing near the margin
above, later conspicuous, somewhat prominent below, veinlets somewhat
conspicuous above, below forming a delicate rather prominent reticulation,
margin subentire or more or less obscurely crenulate, petiolule about 2 mm long,
covered with dense, curly pubescence. Panicles terminal,
subcorymbose, 15 cm long; pedicels 3-4 mm long with short curly
pubescence. Sepals 5, ovate, lanceolate or deltoid, obtuse, 1.25 mm
long, covered with short, curly pubescence without. Petals 5,
linear-oblanceolate, incurving-apiculate, 6.5 mm long, 1.75 mm wide, glabrous,
thickened at the middle, imbricate. Disk fleshy, glabrous, slightly
shorter than the calyx. Stamens 10, alternate ones longer,
filaments 4-5 mm long, narrowed toward the apex, the lower half slightly
broader but scarcely complanate, anthers about 1 mm long, papillate.
Ovary slightly oblong, scarcely 1.5 mm long, glabrous, shortly stipitate; style
3-5 mm long, articulated at the base, glabrous, stigma capitate.
Fruit (immature), 1 cm diam."
Tanaka
(1929a, p. 34) gave a redescription of the material cited by Craib which
adds a few points to Craib's diagnosis. The leaves are clearly
dotted on the upper surface; the terminal inflorescence is small, bearing 10-15
flowers; "flower buds small, oblong-ovoid, about 5 mm long, without glandular
dots, pedicels short, about 6 mm, densely pubescent…ovary cylindric, notched,
continuous with the cylindric style." Tanaka reported that the
fruit clusters are as much as 9 cm wide; the fruits are large, "black, somewhat
4-angled, indistinctly divided by shallow longitudinal grooves"; oil-dots
large, more or less indistinct, pustulate; calyx lobes persistent, with long
soft hairs; "pedicel about 1.5 cm long, continuous to branch of peduncle…about
2 cm long." He found that the fruits, usually about 1.5 cm diam.
may attain 2 cm, and the fresh fruit may measure still more.
Tanaka (1929a, p. 34) stated: "This
species is closely allied with C. [Murraya] koenigii, but
dots on the petals are nearly indistinct, filaments are less subulate, petals
much narrower, and fruit much larger and 4-grooved. The pubescence
of the plant is more pronounced than C. [Murraya]
koenigii, and sometimes the whole plant is covered with white curved hairs,
even…the upper surface of the leaflets." Winit noted on his label on
the type specimen that the plant is decidedly shrubby, blackish with deeply
cracked bark, and has greenish-white, scented flowers.
A specimen from Thailand, collected on December
31, 1928, by Dr. A. F. G. Kerr (No. 9740) has apparently mature, globose, or
slightly depressed-globose fruits, about 1.5 to 1.7 cm in diameter in the dry
state, that do not show any distinct longitudinal grooves but merely very
broad, very shallow depressions between the slightly protuberant locules.
8. Murraya euchrestifolia
Hayata, Icon. Pl. Formos. 6:11. 1916. Clausena euchrestifolia
(Hay.) Kanehira, Formos. Trees 97. 1917; Chalcas euchrestifolia (Hay.)
Tan. Jour. Soc. Trop. Agr. 1:32. 1929. Illus. Kanehira, Formos.
Trees 104. 1917, and ibid. rev. ed. 309, fig. 263.
1936.
Type.—Formosa, Nantôchô, (T.
Itô). Herb. Imp. Univ., Tokyo.
Distribution.—Taiwan.
Tanaka
(1929a, p. 32) described this species as follows: "Shrub; leaflets up to
8, rachis fine, puberulent; leaflets elliptic, acuminate at both ends, shiny
above, largest 9.5 X 3.4 cm, nearly glabrous, lower surface of the midrib
slightly puberulent, margin entire. Inflorescence [a] terminal,
many-flowered cyme, buds small, about 4 X 1.4 mm, black dotting few on the
upper half. Calyx well lobed, about 1.5 mm diam., lobes triangular,
black, glabrous down to the base, margin not pilose; pedicel slender, 4-5 mm
long, pilose with soft, golden, curved pubescence. Petals 5,
oblong-oboval, about 4.5 X 1.5 mm, obtuse at the apex, not much narrowed at the
base, with approximately 10 dots on outside. Stamens 10, one shorter
after the other, [longer and shorter ones alternating], longer ones about 4.5
mm, nearly as long as the petals when fully expanded, filaments linear, flat,
gradually narrowed and acutely pointed; anther small, roundish, slightly
cordate, about 2-3 mm diam. Pistil slightly shorter than the
stamen; ovary oval, constricted at the base toward the broad end; style
cylindric, terete, often slightly curved, stigma obscure, altogether about 3.5
mm long. Berry globose, not apiculate, largest about 1 cm across,
smooth, rather sparingly large pellucid-dotted, dark colored."
Tanaka (1929a, p. 32) stated that this
plant most closely resembles M. crenulata in appearance, but the calyx
lobes are acutely triangular, and the leaflets are not so oblique, lacking
crenulation. In this respect, it approaches M. koenigii more
closely, although the number of leaflets is fewer and they are larger and
glabrous. It differs from both species in having a smaller flower,
an obscure stigma, and a slightly constricted ovary.
9. Murraya crenulata (Turcz.)
Oliv. Jour. Linn. Soc. Bot. 5(2):29. 1861. Glycosmis crenulata
Turcz. Bull. Soc. Nat. Mosc. 31:250. 1858; Micromelum coriaceum
Seem. 1865; Clausena worcesteri Merr. 1910; C. citriodora Merr.
1925.
Type.—Philippines (Cuming, No.
335). Herb., Leningrad.
Distribution.—Java, Philippines, Celebes, New Caledonia.
Tanaka (1929a, p. 30) described this
species as follows: "Shrub; leaves pinnate, composed of about 6-7-paired
leaflets; leaflets medium-sized [3-6 X 1-2 or 3 cm], much oblique, oval or
oboval, acuminate or mucronate at the apex with blunt tip, acute at the base
(roundish in the lower leaflets), very short petiolulate, veins very distinct,
irregular, margin more or less crenulate, semicoriaceous, shining above, drying
black, and midrib always glabrous. Inflorescence a terminal cyme,
many-flowered; flower small, but cylindric, about 5 X 2 mm, apex rounded;
petal narrow, striated, with obscure black dots. Calyx cupulate,
puberulent, lobes broad and rounded, side often imbricate and ciliate on
margin; peduncle slender, tomentose. Stamens free, unequal in
length, filaments long, linear, more or less dilated at the lower half, very
sharp pointed at the apex, anther globose. Pistil nearly as long as
the filaments; ovary ellipsoid, apex not narrowed, rather broad and more or
less ridged, base somewhat narrowed to a cylindric disk; style long, terete,
cylindric, stigma globose. Berry nearly globose and minutely
apiculate, dark-colored."
Tanaka noted that
M. crenulata is allied to M. koenigii but differs from it in
having fewer leaflets, which are thicker, larger, and attached to a glabrous
instead of a pubescent rachis. The fruit is subglobose and minutely
apiculate, whereas in M. koenigii it is short-ellipsoid and
apiculate. M. crenulata is often confused in the field with
Clausena anisum-olens and with Micromelum minutum var.
curranii, but it has long flower buds and long pistils, which are never
found in Clausena or Micromelum.
SPECIES OF UNCERTAIN RELATIONSHIPS
10. Murraya glabra (Guill.) Swing. Jour. Wash. Acad. Sci.
28:532. 1938. Micromelum glabrum Guill. in Lecomte, Not. Syst.
1:216. 1910; Chalcas glabra (Guill.) Tan. 1929. Illus.
Guillaumin, in Lecomte, Fl. Gén. Indo-Chine 1:646, fig. 68(11-12).
1911.
Type.—French Indo-China,
Kien-khé, Tonkin (Bon, No. 2977). Herb. Mus. Hist. Nat.,
Paris.
Distribution.—Vietnam: Tonkin
and Annam.
Guillaumin (1911, p. 650) described
this species as follows: "Small shrub, 1 m high. Branches entirely
glabrous, with very odorous oil glands. Leaves 12-46 cm long;
leaflets 3-9, alternate, glabrous, obovate or oblong-lanceolate, 5-17 X 2-9 cm,
inequilateral and pointed at the base, abruptly acuminate at the tip, very
slightly denticulate; lateral veins, 6-9 pairs, prominent on lower surface;
petiolules cylindric, glabrous, 3-4 mm long; petiole cylindric, glabrous,
glandular. Inflorescences shorter than the leaves, with very short
pubescence; pedicels longer than the flowers, with 2 small bracts at the base or
lower third; flower [buds] 4 mm long, white. Calyx of 4 connate,
indistinct, very shortly ciliate sepals. Petals 4, length 6 mm,
glabrous, lanceolate. Stamens 8, of which 4 are longer; filaments
dilated and pubescent above; anthers attached by the back, oval, with a few
hairs. Disk extremely short. Ovary cylindric, warty,
glabrous; style longer than the ovary, ending in a globose stigma."
Tanaka (1929a, p. 36) in his monograph of
the genus Chalcas (= Murraya) made a few additions and changes to
Guillaumin's description, among others the following: "Leaflets 3-11, generally
5…texture thin, chartaceous…Rachis rather thin [slender?], about 28 cm
long…Flower bud about 6 X 2.5 cm. Ovary…distinctly constricted at
about one-third from the apex…"
This
anomalous species was described as " belonging to Micromelum but was
transferred to Murraya by Tanaka "on account of having a large cyme, long
flower buds with prominent dots and elongated style." The specimens
from Annam have, so Tanaka found, distinctly terminal inflorescences which
bear nearly 100 flowers. The fruit is unknown.
11. Murraya stenocarpa (Drake)
Swing. Jour. Wash. Acad. Sci. 28:532. 1938. Atalantia stenocarpa
Drake, in Morot, Jour. Bot. 6:277. 1892; Glycosmis stenocarpa
(Drake) Guill. 1912; G. bonii Guill. 1910; Chalcas stenocarpa
(Drake) Tan. Bul. Soc. Bot. France 75:711. 1928. Illus.,
Guillaumin, in Lecomte, Fl. Gén. Indo-Chine 1:667, fig. 71(3-6).
1911.
Type.—French Indo-China, Tonkin
(Balansa, No. 1110). Herb. Mus. Hist. Nat.,
Paris.
Distribution.—North Vietnam:
Tonkin.
The original description of
Atalantia stenocarpa by Drake del Castillo may be translated as follows:
"Small tree (up to 1 m high), spineless, very glabrous; leaves subcoriaceous,
shining, elliptical or oblong-lanceolate (leaf blades up to 15 cm long, 6 cm
wide; petiole about 4 cm long), acute at the base, narrowed at the apex, obtuse
or subemarginate, serrate. Cymes axillary,
few-flowered. Flowers small. Calyx lobes 5, short,
ovate, acute. Petals oblong. Stamen filaments free,
ciliate. Ovary oblong, substipitate, with 2 ovules.
Fruit oblong, shortly attenuate at the apex."
A redescription by Guillaumin (1911, p. 671) under the name Atalantia
stenocarpa added a few details "…Leaflets 4-12.5 X 3-5 cm, cuneiform at the
base, attenuate and slightly emarginate at the tip; with 9-10 pairs of lateral
veins, clearly distinct from the veinlets and like them elevated above the
surface below; oil glands easily visible principally on the margins and in the
angles of the teeth; petiole 2-3.5 cm, with a swelling at the base of the leaf
blade; ovary ovoid, borne on a low disk, with 2 locules, each with 1 ovule;
style cylindric-pubescent. Fruits fleshy, reddish, oblong, 15 mm
long, attenuated at the tip."
This anomalous
species, with unifoliolate leaves, was placed in this genus by Tanaka, although
originally it was placed in the genus Atalantia. Guillaumin,
because of this leaf-character, transferred the species to Glycosmis,
but, according to Tanaka, "the plant lacks ferruginous pubescence characteristic
to Glycosmis…The ovary…is glandular and somewhat stalked at the base and
this character agrees with Clausena and not…Atalantia.
The filaments of Clausena, however, are very much attenuated, ending with
a filiform tip, and the anther is extremely long. In the present
species, the filaments are dilated [at the base] and abruptly pointed [at the
tip] and the anthers are not large; moreover they are pubescent, and [have]
pellucid dots on the surface of the petals and calyx lobes similar to
Murraya glabra."
Tanaka (1928b,
p. 711) emphasized the "exact coincidence of the characters of the stamens in
the present species" with those of M. glabra. However, M.
glabra is itself a somewhat anomalous species of Murraya, and both of
these species should be studied more fully.
Subtribe 3.
Merrilliinae: Large-Fruited Remote Citroid Fruit Trees
The subtribe Merrilliinae contains only one genus
with a single species, Merrillia caloxylon native to the Malay Peninsula
and to the adjacent coast of Sumatra. Merrillia differs
widely from all the other genera of the tribe Clauseneae and has a superficial
similarity with the genus Swinglea, one of the aberrant Hard-Shelled
Citroid Fruit Trees belonging to the tribe Citreae.
Swingle follows Engler (1931, p. 211) in placing
Merrillia next to the genus Murraya, which it resembles in general
aspect and in its hard handsome wood, although differing greatly in flower and
fruit characters. The flowers are the largest known in the orange
subfamily but do not open fully (as do those of Citrus).
They are trumpet-shaped, becoming completely pendent during anthesis, with only
the upper third of the petals recurved. The petals composing the
corolla are not only the largest known in the orange subfamily, but they are
also slightly unequal in size and shape, i.e., slightly
zygomorphic. The fruits are as strange as the flowers and have a
thick, radially lacunose, leathery exocarp and seeds covered with abundant,
narrow, flattened somewhat deeply laciniate paleae, characters not found
elsewhere in the subfamily.
This subtribe
seems to have been derived as an offshoot from the ancestors of Murraya
that evolved rapidly in flower and fruit characters while retaining the general
facies of a Murraya of the M. paniculata group of
species.
V. Merrillia Swing.
V. Merrillia Swing. Phil. Jour. Sci. Bot. 13:337.
1918.
Type species.—Murraya
caloxylon Ridl. = Merrillia caloxylon (Ridl.)
Swing.
Distribution.—Malay Peninsula:
southern Thailand, upper Perak; northeastern Sumatra. Cultivated in
Singapore Botanic Gardens.
Common
name.—Merrillia.
Swingle's original
description, translated, reads as follows: "Genus allied to Chaetospermum
[now Swinglea]…from which it differs chiefly in the 5-6-locular ovary
having an irregular lacunose pericarp; the seeds densely scaly, scales
elongate, membranaceous, slightly laciniate; the leaves sessile, pinnate, the
rachis narrowly winged, and the leaflets alternate.
"Unarmed tree; leaves pinnate, sessile or
subsessile; rachis narrowly winged; leaflets alternate, lower ones small, the
next higher ones gradually increasing in size, the terminal ones largest;
petiolule very short; inflorescence axillary, 2-1-flowered; flowers perfect,
rather large, 5-merous; calyx 5-merous, small, lobes triangular-ovate; petals
5, narrow; stamens 10, free, unequal; ovary 5- (rarely 6-) locular, stipitate,
merging into the elongate style, stigma capitate, ovules 8 to 10 in each
locule; fruit subglobose, large pericarp very thick, coriaceous, irregularly
lacunose with cartilaginous walls, segments filled with mucilaginous gum, seeds
numerous in each segment (8-10), lenticular, densely scaly, scales slightly
fimbriate, membranaceous, elongate, hilum carinate, subariloid.
Cotyledons hypogeous in germination, first leaves simple, opposite, broadly
ovate-lanceolate."
The very long,
trumpet-shaped, slightly zygomorphic flowers of Merrillia, often 55 to 60
mm long, are unique in the orange subfamily. The large fruit with
its thick, radially lacunose epicarp and seeds covered with flat, narrow,
laciniate paleae are equally strange. These and other remarkable
flower and fruit characters of Merrillia constitute one of the most
striking examples of rapid divergent evolution to be found among the many that
are seen in the orange subfamily, such as the highly xerophytic Eremocitrus
and the anomalous Monanthocitrus with its horned fruits and spotted
seeds. The extraordinary fact is that Merrillia not only has
flowers, fruits, and seeds which differ very widely from those of the other
genera of the tribe Clauseneae, but fruit characters analogous to those of the
genus Swinglea, an aberrant member of the Hard-Shelled Citroid Fruit
Trees belonging to another tribe.
When Swingle
described Merrillia as a genus in 1918, he thought that it probably
belonged with Swinglea in a special group of the subtribe
Balsamocitrinae, although he directed attention to the unusual characters that
make it "perhaps the most aberrant of the citrus fruits." Swingle
had the good fortune to be able for several consecutive years to study in minute
detail the leaf and flower morphology of a healthy, everbloomingtree of this
plant growing in the citrus greenhouse of the former Bureau of Plant
Industry. As a result of these studies Swingle considered
Merrillia to be a very anomalous member of the tribe Clauseneae that has
probably evolved from a Murraya-like ancestral
form.
Merrillia caloxylon (Ridl.)
Swing. Phil. Jour. Sci. Bot. 13:338. 1918. Murraya caloxylon Ridl.
Jour. Straits Branch Roy. Asiat. Soc. 50:113-14. 1908. Illus.
Swingle, loc. cit. pls. 5, 6; Foxworthy, Malay. For. Rec. 3:169,
unnumbered fig. 1927; fig. 3-10 this work.
Type.—Upper Perak, Kenering, at 500 ft. elevation (Robinson, No.
5548). Herb. Brit. Mus.,
London.
Distribution.—Malay Peninsula:
southern Thailand, Patani Province; Federation of Malaysia: Kedah, Perak,
Selangor, Pahang; northern Sumatra: Asahan.
Common name.—Flowering merrillia (or
ketengah).
The original description of
this species (Ridley, l.c.) follows: "A tree of considerable size, the branches
covered with a pale flaky bark. Leaves 8 inches [20.3 cm] or more
long, with 13 leaflets; rachis flattened and winged, narrow; leaflets 3-3 1/2
inches [7.6-8.8 cm] long or less by 1 1/2 inches wide [3.8 cm], alternate,
oblanceolate, obtusely acuminate with a triangular base, minutely petiolate,
inequilateral, thin, bright deep green. Flowers pale
yellowish-green, several together in small panicles, in the upper axils of a
branch, about an inch [2.5 cm] long. Sepals connate, ovate-acute,
1/10 inch [2.5 mm] long. Petals and stamens not seen.
Ovary stalked, hairy, style rather stout, hairy, stigma
capitulate. Fruit oblong, rounded at both ends, 4 inches [10.2 cm]
long and three inches [7.6 cm] in diameter, the pericarp dotted and warty,
greenish, eventually becoming yellow, half an inch [12.7 mm] thick, lemon
yellow inside, full of long resin cells narrowed at the mouth and dilated
below, cells 5, with rather thick tough walls, pulp of transparent flattened
sticky fibers, olive green in colour and tasteless. Seeds numerous,
about 5 in a section, ovate-flattened, half an inch [12.7 mm] long, 1/8 inch [3
mm] thick, olive gray."
Open flowers of this
species were not seen by Ridley. As Swingle reported in the
original description of Merrillia, petals from flowers of a tree in the
Singapore Botanic Gardens grown from seed from Patani Province, in extreme
southern Thailand, were "35-40 X 5-10 mm, bluntly pointed at the apex and
narrowed gradually to the base." A small tree grown in the citrus
greenhouse at Washington, from seeds of this tree in the Singapore Botanic
Gardens, bore flowers that were even larger, having petals 4.5-5.5 cm long and
8-11 mm wide. The flowers were pendent and never opened fully,
having the imbricate petals curving outward at the tips and making a
trumpet-shaped flower, greenish-white on the outside and white
within. No other species of the orange sub-family has flowers of
this type. Swingle directed attention (1918, p. 339, pl. 5) to the
curious pair of nearly opposite, small, broad, and often suborbicular leaflets
which are borne very near the base of the rachis, much nearer than in any other
pinnate-leaved species in the orange subfamily (fig. 3-10).
This remarkable
species, called ketenggah in the Malay Peninsula, resembles Murraya
paniculata in its general habit of growth and in its leaves, and even in the
texture and color of its wood. These two species are sometimes
confused by the natives under the name kemuning, which properly applies
to Murraya paniculata. The flowers of Merrillia are
entirely different, and its fruits are still more unlike those of
Murraya, so it stands out taxonomically as a strikingly different plant
obviously belonging to a distinct genus.
This
tree well deserves to be grown in greenhouses or out of doors in warm
subtropical climates as an ornamental. When a small tree, 3 to 3.5
m high, it flowers abundantly at frequent intervals, almost all the year
round. Its curious, pendent, trumpet-shaped flowers, although not
conspicuously colored, are attractive because they are produced in such
abundance and are pleasantly fragrant.
In his
notes following the original description of Murraya caloxylon Ridley
stated that it "is famous in the Malay Peninsula for its beautiful wood," which
"is of a bright yellow color ornamented with dark brown streaks and stains,
fairly hard in texture and taking a good polish." It is said to be
highly prized by the Malayans for making walking sticks, kris handles, and
other small objects. A British official made beautiful furniture,
boxes, etc., out of the wood. Foxworthy (1927, p. 170) considered
it a slow-growing species and thought that it would require about thirty years
to reach commercial size. It is so rare in the wild state that any
considerable supply of wood would have to come from cultivated plants.
This species, or one very closely related to it,
has been found in northeastern Sumatra directly across the Malacca Strait from
its habitat in the Malay Peninsula. Mr. B. A. Krukoff, in
November-December, 1932, collected good fruiting material of this tree "in
primary forests on ‘red’ soil" at Hoeta Padang, near Asahan, northeastern
Sumatra (Krukoff, No. 4319, in Herb. Arnold Arbor.). Krukoff noted
on the label "tree 110 ft. [33.6 m] high," i.e., nearly twice the height yet
reported for M. caloxylon growing in the Malay Peninsula! An
earlier collection made at the same place in December, 1930, by Krukoff was
from a small tree only 15 cm in diameter. Mr. C. G. G. J. van
Steenis collected in 1932 a leafy branch of Merrillia at 120 m altitude
on Mount Maligas, Simeloengoen, eastern Sumatra (van Steenis, No. b.b. 20415,
in Herb. Arnold Arbor.). No other tree in the orange subfamily has
been found to attain the height reported by Krukoff. If such giant
trees produce wood as beautiful as the smaller form growing in the Malay
Peninsula, it would be worth while to test them under
cultivation.
TRIBE II. CITREAE: CITRUS
AND CITROID FRUIT TREES
The tribe Citreae
includes Citrus and all of its near relatives as well as many more remote
relatives that have not yet been hybridized with Citrus or grafted on
it. The Citreae may be described as follows:
Small trees or woody, clambering lianas almost
always with single or paired axillary spines at the nodes of the twigs; leaves
persistent (except in three monotypic genera, Poncirus, Aegle, and
Feronia), usually simple or 1-3-foliolate (usually pinnately 3-foliolate
with a short petiole and the middle leaf larger, but in Luvunga palmately
3-foliolate with a very long petiole) or sometimes odd-pinnate with strictly
opposite pinnae; petioles often winged; flowers in axillary clusters, or in
terminal or axillary panicles or corymbs; flowers white or greenish, usually
fragrant; calyx with 3-5 sepals or lobes; corolla imbricate in aestivation,
with 3-5 (rarely 6) petals; stamens 2-4 or more times as many as the petals;
ovary 2-18-locular, ovules 1-18 (usually 1-12) in each locule (22 or more in
Burkillanthus); fruits usually orange- or lemon-like in external
appearance, often small, sometimes small red or nearly black juicy berries; in
the subtribe Citrinae the locules contain pulp-vesicles filled with juice but
in the other two subtribes (Triphasiinae and Balsamocitrinae) the locules lack
pulp-vesicles, but are usually filled with a mucilaginous gum; seeds with one
or several embryos; if with several, only one (rarely two) is a true embryos,
the others being false embryos that develop from nucellar buds that grow into
the embryo sac; germination hypogeous or epigeous.
Almost all the species develop single or paired
spines in the axils of the leaves of vigorous shoots, especially on young
trees. The leaves are simple, unifoliolate, or trifoliolate in most
of the genera, but a few genera, Hesperethusa, Feroniella, and
most of the species of Citropsis have odd-pinnate leaves with the
leaflets attached to the rachis in strictly opposite pairs (very different from
the alternately attached leaflets of the tribe Clauseneae). Such
odd-pinnate leaves often have the petioles and the segments of the rachis
between the pairs of leaflets broadly winged. The rachis is
articulated at the point of attachment of each pair of pinnae and these
articulated segments fall apart when the leaf finally falls off.
The leaves of Luvunga are palmately trifoliolate with very long
petioles and pulvini at the bases of the petiolules and of the
petioles. Three genera, Poncirus, Aegle, and
Feronia, have deciduous leaves; all the other genera have persistent
(evergreen) leaves. The species with trifoliolate and unifoliolate
leaves usually have winged petioles, whereas the species with simple leaves
usually have wingless petioles, although the margins of the leaf blade are
sometimes decurrent on the petiole, for part of its length. The
ovary has 2 to 18 or 20 locules, each with 1 to 18 ovules (22 or more in
Burkillanthus) in each locule. The ovaries with 2 to 5
locules usually have one or two ovules in each locule (except in
Burkillanthus); those with 6 to 18 locules usually have 4 to 16 or even
18 ovules in each locule. The fruits are often small, but they are
large or very large in Citrus and in Burkillanthus and in several
of the genera of the subtribe Balsamocitrinae.
In the course of an investigation of the vascular traces of all obtainable
genera of the orange subfamily, Tillson (1938, pp. 12, 25; Tillson and Bamford,
1938, pp. 785, 789, 791) found that most of the genera of the tribe Citreae
studied (21 out of 24) showed fusion of the bundles which arise from the axis
and extend to the sepals and petals; "the sepal midribs carry out the lateral
petal bundles fused to them, and in some species the lateral sepal bundles are
fused in varying degrees to the petal midribs." The three closely related genera Aegle, Afraegle, and
Aeglopsis are without fusions of either kind. None of the
many species belonging to the five genera of the tribe Clauseneae showed
fusions of sepal and petal bundles.
The tribe
Citreae is distributed in southeastern Asia and the Monsoon region to the
southeast as far as eastern Australia, New Caledonia, New Guinea, and the
Melanesian islands; also in tropical (and subtropical) mountainous regions in
eastern, central, and western Africa.
The
tribe Citreae contains, as here treated, twenty-eight genera with 124 species,
which fall very naturally into three subtribes. A key to the subtribes of Citreae is presented.
Subtribe 1.
Triphasiinae: Minor Citroid Fruit Trees
Small trees or woody climbers, usually with single or paired spines in the
axils of the leaves, but often spineless; leaves pinnately 3-foliolate
(palmately 3-foliolate in Luvunga), often 1-foliolate or even simple;
fruits small, often orange- or lemon-like in appearance, rarely small red or
black juicy berries; fruit locules without pulp-vesicles, but usually filled
with mucilaginous gum; ovary subsessile or more or less stalked, borne on a
disk or on a longer cylindrical gynophore with 2-5 (rarely 1) or even 6 or 7
locules, and 1-6 or 8 ovules in each locule; fruits with gland-dotted peel,
sometimes soft and berry-like, usually firm and often resembling miniature
oranges or lemons but always lacking pulp-vesicles.
Most of the eight genera of the subtribe
Triphasiinae contain species with one or two axillary spines.
However, all four species of the genus Oxanthera are thornless and
Wenzelia has six thornless species (also two with single spines and one
with paired spines). All the species of this subtribe have
persistent evergreen leaves but they vary greatly in the different genera:
Wenzelia, Monanthocitrus, Merope, and Pamburus have
simple leaves with very short, wingless, non-articulated petioles; Triphasia
has pinnately trifoliolate or simple leaves; Luvunga has palmately
trifoliolate leaves with the leaflets attached by pulvinoid petiolules to a
long petiole with a pulvinus at the base; Paramignya has unifoliolate
leaves with wingless petioles usually pulvinoid at the junction with the blade
(the two last-named genera have petioles articulated with the leaf blade);
Oxanthera has three species with unifoliolate leaves with articulated
petioles and one simple-leaved species with nonarticulated petioles.
The eight genera of this subtribe are all native
to an area partly superimposed on that in which the True Citrus Fruit Trees
occur, but this area, instead of being barrel-shaped, is rather a long,
obtuse-angled triangle with its acute apex cut off, having a land base in Asia
extending about 4,600 kilometers from western India to southeastern China and
Hainan Island and reaching some 10,000 to 11,000 kilometers to the southeast
into Polynesia. This area falls mostly in the Monsoon
region. Its middle portion is about 3,000 kilometers wide from
Sumatra to the Philippines and its blunted southeastern apex, which is bounded
by New Caledonia and the Fiji Islands, is about 1,800 kilometers wide.
Of the eight genera included in this subtribe,
only two, Oxanthera and Monanthocitrus, have a very restricted
range, Oxanthera being found only at the very point of the triangle, in
New Caledonia (and near-by Art Island), and Monanthocitrus in
west-central New Guinea. Merope (which has leaves and
petioles resembling Wenzelia) occurs in brackish coastal swamps from
northwestern New Guinea and Amboyna Island to the southwestern Philippines,
Java, Malay Peninsula, Burma, and Bengal. Wenzelia,
Merope, and Monanthocitrus all include New Guinea in their range
and may possibly have originated there. These three genera and the
related genus Oxanthera are the only ones in the subtribe with more than
two (4 to 8) ovules in each locule. The other four genera (all with
one or two ovules in each locule) seem to have originated in southeastern Asia
and to have spread a greater or less distance into the East Indian
Archipelago: Triphasia and Luvunga range clear through to New
Guinea; Paramignya ranges to Sumatra and the Philippines; whereas
Pamburus has spread only as far as Sumatra, Java, and Borneo.
The subtribe Triphasiinae contains three fairly
distinct groups of genera, although most of the genera included differ
strikingly from one another. Furthermore, within a given genus, the
species are far from uniform, many of them differing in characters usually
considered to be of high taxonomic value. The genus
Oxanthera, for example, has only four species, but they run the whole
gamut of ovarial morphology: O. aurantium has a hypomerous ovary with two
locules; O. fragrans has an isomerous ovary with 5 to 6 locules; whereas
O. neo-caledonica has a hypermerous ovary with seven locules.
Several other genera in the subtribe have aberrant species and doubtless others
will be discovered in the future. No other subtribe in the whole
orange subfamily gives such strong evidence of containing species that have
been for ages and probably still are rapidly evolving into new forms, new
species, and new genera.
The three groups of
genera within the subtribe Triphasiinae are indicated in the key to the genera and are described briefly in the
text. The Wenzelia group has four genera, all with more than
two ovules (4 to 8, usually 6) in each ovary locule and long, simple or
unifoliolate leaves, with very short nonarticulated petioles except in
Oxanthera. The Triphasia group contains two genera,
the species of which are all small trees or shrubs with short nonarticulate
petioles. Two species of Triphasia have simple leaves and one
has trifoliolate leaves; the other genus in the group, Pamburus, has
simple, semicoriaceous thick leaves. The Luvunga group has
two genera, both woody lianas that clamber over trees by means of recurved or
hooked spines, with small or medium-sized ovate or subglobose fruits that look
like miniature limes.
The remote ancestors of
our cultivated citrus trees doubtless were very like some of the plants now
found in this subtribe. Doubtless several tens of millions of years
have elapsed, however, since the subtribe Citrinae arose from some ancestral
species similar to those now classed in Triphasiinae.
The forty-six species known in Triphasiinae make
it the second largest of the three subtribes in the tribe Citreae.
It contains the nearer wild relatives of our Citrus Fruit Trees, all of which
should be introduced into culture, studied, tested and utilized as rootstocks
or in other ways by citrus growers and citrus experts the world
over. It is noteworthy that no fewer than twenty-nine species
(nearly three fifths the total number!) have been described as new and also
that three of the eight genera have been established since 1900. As
the rich flora of the Monsoon region is more and more actively assembled by
competent collectors and studied by expert taxonomists, there is good reason to
expect that many more species and doubtless some new genera will be discovered
in the future.
GROUP A. THE WENZELIA
GROUP
The Wenzelia group consists of four
genera: Wenzelia, Monanthocitrus, Oxanthera, and
Merope. The first three have six ovules in each ovary locule
and the last one has four ovules to each locule. The leaves of the
plants of all four genera are simple or unifoliolate.
Wenzelia, Merope, and Monanthocitrus all occur in New
Guinea and are closely related genera. Oxanthera, found in
New Caledonia, in spite of its long isolation in both time and space, is still
obviously related to Wenzelia. Merope has leaves very like
those of Wenzelia and Monanthocitrus, but each ovary locule, so
far as known, contains four (or by abortion two?) ovules and its curious
flattened seeds, the largest known in the orange subfamily, are of unusual
shape, being more or less caudate. Merope is doubtless an
early offshoot from an ancestral form of the Wenzelia group.
Wenzelia and Monanthocitrus differ greatly from all the other
Minor Citroid Fruit Trees in having large leaves with extremely short petioles
not articulate with the leaf blade.
In
Oxanthera the leaves, the ovaries, and the stamens have varied from the
primitive pattern found in Wenzelia and
Monanthocitrus. In Merope the seeds have evolved along
new lines; in Monanthocitrus the seeds have continued to evolve along
lines shown by certain flat-seeded species of Wenzelia until they now
show characters found nowhere else in the orange subfamily.
Clymenia, a True Citrus Fruit Tree, has leaves very like those of
Wenzelia but the fruit shows stalkless pulp-vesicles that pave the way to
Citrus.
All the plants of the
Wenzelia group should be studied attentively to throw all the light
possible on the origin of Citrus.
VI. Wenzelia Merr.
VI. Wenzelia Merr. Phil. Jour. Sci. Bot. 10:272.
1915.
Type species.—Wenzelia
brevipes Merr., from Leyte Island, Philippines (Wenzel, No.
998). Herb. Bur. Sci., Manila.
Distribution.—Southern Philippines, New Guinea, Bougainville island, Solomon
Islands, Fiji Islands.
Common
name.—Wenzelia.
Shrubs with unarmed twigs
or with single or paired spines in the leaf axils; leaves large (7-28 cm long),
oblong or lanceolate; petioles very short (2-5 mm long), wingless but channeled
above, not clearly articulated with the leaf blade; flowers often very large;
solitary in the axils of the leaves, white; calyx 5-lobed; petals 5,
oblong-lanceolate or ovate, obtuse or subacute, glandular-punctate; stamens 10,
free, anthers long; ovary oblong-ellipsoid, 4-5-locular with 6 (or sometimes 8)
ovules in each locule, narrowed below into an annular or shallow cupulate disk;
fruit obovoid or ellipsoid, 3-5 cm long, with coriaceous, glandular pericarp,
segments containing mucilage without pulp-vesicles.
The Wenzelia genus is very different from
all the other Minor Citroid Fruit Trees of the subtribe Triphasiinae except
Monanthocitrus in having large, or very large, simple leaves, with very
short wingless petioles not articulated with the leaf blade, and in having six
or eight ovules in each locule of the ovary. The fruits are
ellipsoid and the locules contain several seeds (up to six or eight) immersed
in a mucilaginous material in young fruits.
Nine species of Wenzelia are known, six from New Guinea, one from the
southern islands of the Philippines, one from the Solomons, and one (the
easternmost large-seeded member of the orange subfamily) from Fiji.
Seven of the species fall into two groups constituting the two subgenera
described below, leaving two species still inadequately known and of uncertain
relationships.
Wenzelia is the largest
and most widely distributed of the six genera limited to the southern part of
the Monsoon region, Melanesia, and the Fiji Archipelago, the southeastern area
of distribution of the orange subfamily. Three of these six genera,
Clymenia (Bismarck Archipelago), Monanthocitrus (New Guinea), and
Eremocitrus (northeastern Australia), are monotypic; Oxanthera
(New Caledonia) has four species; Microcitrus (northeastern Australia and
southern New Guinea) has six species; Wenzelia (the Philippines, New
Guinea, the Solomons, and Fiji) has nine species!
A striking proof of the recent rapid advance of
our knowledge of Citrus relatives is provided by the fact that all the
nine species now constituting the genus Wenzelia have been described
since 1900. The first species was published in 1901, the second in
1910, the third in 1915 (when the genus Wenzelia was established), a
fourth in 1918, and the other five were described early in 1940 (see Swingle,
1940a).
It is very probable that still
other species of Wenzelia will be found when New Guinea, the Spice
Islands (the Moluccas), and the Melanesian islands to the east of New Guinea
are explored more thoroughly. Keys to the subgenera and species of Wenzelia are
presented.
Subgenus Wenzelia
Subgenus
Euwenzelia Swing. in Webber & Batchelor, Citrus Indus. 1:216.
1943.
This subgenus includes Wenzelia
brevipes, W. tenuifolia,and W. kambarae. These
species occupy the two extremes of the area of distribution of the genus, as
well as a midway location in New Guinea: W. brevipes (the type of the
subgenus) is native to the southern islands of the Philippines, W.
kambarae to the Lau Group of the Fijis, and W. tenuifolia to
southeastern New Guinea.
These species
probably resemble the ancestral type of the subgenus Wenzelia, whereas
those of the subgenus Papualimo represent a more highly specialized type
of later origin.
1. Wenzelia
brevipes Merr. Phil. Jour. Sci. Bot. 10:272.
1915.
Type.—Leyte Island, Buena Vista
near Jaro, Philippines (Wenzel, No. 998). Herb. Bur. Sci.,
Manila.
Distribution.—Southeastern
Philippines: Leyte, Samar, and Bohol islands and eastern Mindanao
Island.
Common name.—Philippine
wenzelia.
Merrill’s original extended
description in English, following the Latin diagnosis, reads as follows: "An
erect, glabrous, unarmed shrub about 1 m high, the trunk about 3 cm in
diameter. Branchlets slender, terete, greenish-olivaceous when
dry. Leaves alternate, simple, presenting no indication of a joint
between the lamina and the petiole, firmly chartaceous to subcoriaceous,
oblong, pale when dry, shining, 14 to 25 cm long, 5.5 to 6.5 cm wide,
distinctly glandular-punctate, entire, apex acuminate, base obtuse to rounded;
lateral nerves about 10 on each side of the midrib, prominent on the lower
surface, arched-anastomosing, the primary reticulations lax, prominent;
petioles 2 to 4 mm long. Flowers white, fragrant, axillary,
solitary, their pedicels about 1 cm long with 2 basal, oblong-ovate, 2 mm long
bracteoles. Calyx 5 to 6 mm long, cup-shaped or somewhat
campanulate, 5-lobed, glandular-punctate, the lobes very broadly ovate,
rounded, about 2 mm long, wider than long. Petals 5, imbricate, up
to 3.4 cm long and at least 8 mm wide, oblong-oblanceolate, obtuse, narrowed
below, prominently glandular-punctate. Stamens 10, free; filaments
linear, 1.4 cm long, equal, rather slender, cylindric; anthers oblong,
basifixed, 2-celled, about 3.8 mm long. Disk somewhat cup-shaped,
1.5 mm in diameter, forming a short gynophore. Ovary
oblong-ellipsoid, glandular, about 4 mm long and 2.5 mm in diameter, 4- or
5-celled, narrowed below into the 2 to 3 mm long stalk, and above into the
cylindric style, which is about 9 mm long, continuous with the ovary, ultimately
deciduous; stigma subcapitate, very obscurely lobed; ovules 6 in each cell,
2-seriate, axile. Fruits obovoid, 3-4 cm long, base narrowed, the
pericarp coriaceous-fleshy, glandular; seeds few, apparently 3 or 4 in each
fruit, the cotyledons thick, fleshy, oblong-obovoid, about 1.5 cm long and 1 cm
wide."
Some of the later collections show
broader leaves than those of the type specimen described by
Merrill. For example, a specimen from Bohol, collected by M. Ramos
(No. 43189), has leaves 13.0 to 17.5 by 6.5 to 8.5 cm, acuminate at the tip,
broadly rounded or even slightly cordate at the base, petioles very short, 4 to
5 mm long, 2 to 2.5 mm in diameter, more or less longitudinally rugose, not
articulated with the leaf blades; flower buds single, axillary, 15 by 4 to 5 mm,
pedicel 1 cm long, slender (0.6 to 0.8 mm diam. at base, 1 to 1.3 mm diam. at
tip).
1a. Wenzelia
brevipes var. alabatensis Swing. Jour. Arnold Arbor. 21:16.
1940.
Type.—Philippines, Alabat Island
(Ramos and Edaño, No. 48054, Bur. Sci.). Herb. Arnold Arbor., Harv.
Univ., Cambridge.
Distribution.—Known
only from type locality.
Leaves narrow
(22-28.5 X 5-6.6 cm), much narrower than those of the species; oil glands less
conspicuous, especially on lower surface.
This variety occupies the northernmost point of the range of the species W.
brevipes. Like the species itself and like all other species of
the genus Wenzelia, it is native in an island on the western boundary of
the Pacific Ocean. There is some variation in the shape of the
leaves in the species of the genus Wenzelia, but no other species shows
so extreme a variation as that represented by this variety with its extremely
long and narrow leaves, also peculiar in their less conspicuous oil
glands. It may prove to be an extreme form connected with the
typical form of the species by intergradations.
2. Wenzelia tenuifolia Swing.
Jour. Arnold Arbor. 21:14. 1940.
Type.—Southeastern New Guinea, Papua, Boridi, forests, alt. about 1,220 (or
3,353) meters, "c[irca] 4,000 [or 11,000] feet" (C. E. Carr, No. 14881, 2
branches, 1 fruit, February 1, 1935). Herb. Bot. Mus.,
Berlin-Dahlem; photographs and serial microtome sections, Herb. Natl. Arbor.,
Washington, D.C.
Distribution.—Known
only from the type locality.
A spineless
shrub, 2 m high; ultimate branches slender, 1.1-1.8 mm in diam., internodes 2-3
(rarely 5) cm long; leaves very thin, brittle when dry, more or less broadly
elliptical 10-13 cm long including petiole, 5-7.2 cm wide, apex bluntly
rounded and often imperfectly developed, the very tip being stunted and more
or less torn, sometimes more or less irregularly emarginate, broadly cuneate at
the base, margins entire, lateral veins, 10-12 pairs, arising at angles of
65°-80° with the midrib, petioles 4-6 X 1-1.1 mm; fruit ovate, 3.5 cm long, 3
cm broad, 5-loculate, radial locule walls thin, 1-1.5 mm thick but expanding
into a thick rib of tissue running longitudinally in the fruit, more or less
triangular in cross section, 4-7 mm in radial thickness, dorsal locule walls
only 1-2 mm thick midway between the radial walls; seeds dull brown, ovate,
compressed, 17-19 X 12-15 X 3.5-5 mm, more or less wedge-shaped, with a
faintly, more or less concentrically marked and corrugated edge, 1-2.2 mm wide
and 1-1.5 mm thick, on one side of the seed, with faint markings 0.2-0.4 mm
apart, 1-4 in a locule, often occurring in closely approximated pairs; embryo
monoembryonic.
This species was known to
Swingle only from the rather scanty type specimen, but it is so evidently
distinct from all the other known species that he had no hesitation in making
it a new species. It is somewhat of a connecting link between the
two subgenera Wenzelia and Papualimo but apparently falls in
Wenzelia, as the seeds lack the paper-thin, more or less torn membrane
along the free margins of the seeds found in the species of the subgenus
Papualimo.
The fruits of W.
tenuifolia are circular in cross section and show five more or less
triangular peripheral ribs at the distal ends of the radial locule walls, which
fill the spaces between the strongly bulging dorsal locule walls.
In striking contrast to this species, W. archboldiana has fruits which
are bluntly star-shaped in cross section because of the absence of any tissues
filling the interlocular furrows.
The
collector’s label on the type specimen is a carbon copy (the original label was
doubtless used for another specimen of the plant). The altitude
given on the label apparently reads "c. 11,000 feet" but may possibly be "c.
4,000 feet." If the altitude is 4,000 feet (1,220 meters), it
would be far above the usual upper limit for the aurantioid plants in New
Guinea (about 300 meters, or 984 feet). If the altitude is 11,000
feet (3,353 meters), it would equal the highest altitude as yet reported for
any rutaceous plant from New Guinea.3
As all the other species of Wenzelia grow
at low altitudes in tropical regions, it will be of interest to learn more about
the distribution of W. tenuifolia. Perhaps its curious
unusually thin leaves may be adapted for growth in very humid locations, such
as occur in the cloud belts of mountains.
3. Wenzelia kambarae Swing. Jour. Arnold Arbor. 21:12.
1940. Atalantia sp. Albert C. Smith, Fijian Plant Studies Bul.
141:77. 1936. Illus. Swing., loc. cit. pl. 3, figs.
6-8.
Type.—Fiji Archipelago, Lau Group,
Kambara Island, limestone formation, alt. 10-100 m (A. C. Smith, No.
1265). Herb. New York Bot.
Gard.
Distribution.—Fiji Archipelago:
Lau Group, Kambara Island (A. C. Smith, Nos. 1265 and 1293), and Naiau Island
(Zimmerman). Herb. Natl. Arbor., Washington,
D.C.
Common name.—Fiji wenzelia (or
moli moli).
A spineless shrub or
slender, small tree, 4-7 m high, trunk 4-8 cm diam., twig slender, 1-3 mm diam.,
glabrous, at first green but soon turning light brown and showing numerous
irregular, more or less anastomosing, longitudinal, very narrow ridges; leaves
thin, chartaceous, oblong to elongate-elliptical, varying considerably in size
and shape, 6-21 X 3-8.5 cm, bluntly pointed or rounded at apex, broadly cuneate
or rounded at base, margins entire or irregularly and shallowly crenate
(Zimmerman's Naiau material), lateral veins very distinct on under surface,
less distinct above, 8-13 on each side arising at rather widely varying angles
(60°-80°) with the midrib, branching dichotomously at 3/5-4/5 of the distance
to the margin, petioles 5-7 X 1.5-2 mm, tapering into the leaf blade with a
very shallow channel on the upper side formed by the decurrent leaf margins,
petioles on old leaves often brownish and irregularly ridged or rugose, as is
the midrib on the lower surface; flower buds (coll. by Zimmerman, Naiau Island,
very young) 2.5 X 2 mm, borne on slender pedicels 8-9 mm long, bearing several
minute pointed bracts on the lower portion; fruits orange colored, 3-4 cm diam.,
3- (or 4-?) locular with a gland-dotted but smooth peel, borne on pedicels
14-16 X 1.8 mm, with a cylindrical carpophore 1.2-2.5 mm long between the
persistent calyx and the base of the fruit; seeds very large, ovoid, 18-25 X
12-15 mm, testa smooth, shiny, cream-colored, embryos 1 or 2 (sometimes more?),
greenish-colored.
This species is the
easternmost large-seeded Citrus relative that occurs on any Pacific
island. Its seeds are so large that it is improbable that they
would have been carried to Fiji by birds, especially since the fruits contain no
pulp and little if any mucilaginous matter. It is highly probable
that the moli moli reached the Fiji Archipelago from the center of origin
of the genus Wenzelia (probably New Guinea) by slowly spreading eastward
over dry land, a migration that probably required millions of years to
accomplish. Its nearest known relative is W. melanesica,
native to Bougainville Island in the Solomons, about 1,800 kilometers to the
west-northwest. That dry land once extended from the west as far
east as Fiji in the early Tertiary epoch is considered certain by some
geologists ((Brock, 1924, p. 75).
Dr. Edward
C. Zimmerman, who obtained flower buds of this plant by "shooting" the
dangerous surf on the atoll surrounding Naiau Island, thus making
identification possible, wrote Swingle that the natives of the Lau Group know
the plant well by the name moli moli; furthermore, they told him that
there were pink and red flowers, perhaps in different stages of development (all
other Wenzelia flowers so far observed are white). The trees
on Naiau Island were taller (5 to 7 m) than those reported from Kambara Island
(3 m), but the leaves of the Naiau plants were much smaller (6 to 10 by 2.5 to
5 cm) than those of one of the collections (A. C. Smith, No. 1265) from Kambara
(11.5 to 21 by 4 to 8.5 cm). However, another collection from
Kambara Island (A. C. Smith, No. 1293) shows smaller leaves (8 to 12 by 2.5 to
5 cm). Probably W. kambarae is a variable species showing
many forms.
Subgenus Papualimo
The subgenus
Papualimo includes four species, W. platysperma (the type of the
subgenus) W. paludosa, W. melanesica, and W. archboldiana,
all native to New Guinea or the Solomon Islands to the eastward.
Wenzelia paludosa (Lauterb.) Tan.,
which has very thin seeds (only 1 to 2 mm thick), almost certainly falls in the
subgenus Papualimo, although nothing is known about the margination of
the seeds, a striking character of the other species of this subgenus.
The species constituting the subgenus
Papualimo represents a striking new evolutionary tendency as shown in the
development of thin seeds with very thin, laciniate membranous margins—seeds
that differ widely from the ancestral type of the genus Wenzelia and from
all the other genera of the orange subfamily. This evolutionary
tendency has apparently reached its highest development in the very strange
monotypic genus Monanthocitrus, also native to New Guinea, which has
still thinner, still more elaborately margined seeds. The seeds are
also spotted, unlike those of any other species known in the orange
subfamily.
4. Wenzelia
platysperma Swing. Jour. Arnold Arbor. 21:11. 1940. Illus.
Swing. loc. cit. pl. 3, figs. 4, 5.
Type.—New Guinea, Papua, Palmer River, 2 miles below junction of Black
River, alt. 100 m, flood-plain forest undergrowth (L. J. Brass, No. 7111,
branch with fruit), Herb. Arnold Arbor., Harv. Univ., Cambridge; also found in
the ridge forest undergrowth at the same locality (Brass, No. 7025, branch with
fruit), Herb. Arnold Arbor.
Distribution.—Known only from the two above-cited collections, made very
close together.
A thornless, loosely branched,
small tree, 3 m high; twigs slender, 1-2 mm diam., internodes 2-3 cm long,
often bent slightly at each node alternately to the right and left, glabrous
or very minutely and sparingly pubescent; leaves chartaceous,
elongate-elliptical, 10-20 X 2.5-5.5 cm, acuminate or caudate at apex,
cuneate, or broadly rounded at base, margins irregularly crenulate (and also
denticulate in Brass No. 7025), lateral veins 12-14 on each side, arising at
angles of 70°-80° with the midrib, branching dichotomously at 1/4-3/4 of the
distance to the margin, petioles very short, 3-4 X 1.4-1.8 mm (1/40-1/60 of the
length of the leaf blade), with a shallow channel on the upper side; flowers
not seen, calyx persistent, subtending the fruit, deeply lobed, lobes 2 X 2 mm,
broadly rounded, minutely puberulent (glabrous and apparently slightly fleshy
in Brass No. 7025); fruits subglobose or ovate, 4-4.8 X 3.5 cm, 4-locular (?),
reddish or pink, borne singly on a slender pedicel 10-12 mm long, arising in
the axil of the uppermost leaf; seeds flattened, light brown, 12-15 X 7-9 X 2-3
mm, testa rugose on edges or else tapering into irregularly shaped extensions
with somewhat torn margins, monoembryonic, cotyledons greenish.
This remarkable species is known only from two
single fruiting twigs. Both collections show zigzag slender twigs
each with a single fruit borne in the axil of the uppermost leaf.
The seeds are even flatter than those of W. archboldiana, but otherwise
they are much the same. The globose or ovate, pink or reddish,
gland-dotted, aromatic fruits hang down at the ends of the fruiting
branches. There is no trace of pulp or mucilage between the crowded
seeds that lie lengthwise in the carpels.
5. Wenzelia paludosa (Lauterb.) Tan. Jour. Arnold Arbor.
9:139 1928. Citrus paludosa Lauterb. Bot. Jahrb. 55:263.
1918.
Type.—Northeastern New Guinea, in
swampy primeval forest with sago palms and rattan at mouth of Sepik River
(Ledermann, Nos. 7173 [flowers] and 7507a [fruits]). Herb.
Bot. Mus., Berlin-Dahlem.
Distribution.—Northeastern New Guinea: known only from the Sepik River.
The original description of Lauterbach,
translated, reads as follows: "Shrub or small tree, with smooth, slender,
terete branches, and stout solitary spines; petioles short, caniculate above;
leaves lanceolate, gradually narrowly acuminate, acumen obtuse, emarginate [at
the tip], slightly rounded or subacute at base, chartaceous, glabrous on both
sides, opaque, dull green when dry, 12-14 lateral veins, oblique, dividing near
the margin, many [of them] arched-anastomosing, prominent above when dry like
the midrib, margins undulate-crenate, minutely subdenticulate; flowers
axillary, solitary; calyx 5-lobed, lobes ovate, rounded, ciliate; petals 5,
ovate, subacute, glandular on outer side; stamens 10, filaments free, anthers
linear; disk annular; ovary flask-shaped, locules many-seeded; style thick,
stigma 5-lobed; fruit ovate, subacuminate, exocarp moderately thick,
glandular, pulp scarcity visible; seeds up to 6 in each locule, irregularly
triangular or quadrangular, much flattened, testa white, coriaceous, slightly
rugose.
"Shrub, 1-2 m high, or small tree,
with light gray bark. Branches 2-3 mm thick; petioles 2-5 mm;
leaves 7-14 cm long, 2-5 cm wide; spines 10-12 mm; flower pedicel 6
mm. There are only 2 not fully developed [flower] buds that
measure 3 mm. Fruit pedicel 10 mm; the ripe fruit, which smells
like an orange, is 4 cm long by 3 cm diam.; the seeds 10-12 mm long, 1-2 mm
thick."
A later collection (Ledermann, No.
12252) was made at a hill camp on the Sepik River. The type
specimen showing fruits (Ledermann, No. 7507a) has pedicels of the young
fruits 10 to 12 mm long and unusually slender for a species of Wenzelia,
being only about 0.5 mm in diameter in the middle, tapering to about 1 mm at
the base and also at the top where it bears the calyx without any gradual
expansion into the funnel-shaped base of the calyx as in W.
melanesica. The young fruit is ellipsoid, 14 by 5 mm, narrowed
into a slender stalk at the base, 3 mm long by 1.5 mm thick, and attenuate at
the apex into a beak about 2 mm long and 1 mm wide. The seeds are
flattened, as are those of W. platysperma and W. archboldiana
from the eastern half of New Guinea and W. melanesica from the
Solomons, but unfortunately it is not known whether they are margined or not,
and if so with what kind of a margin. The petioles are very short,
3 to 4 mm long and 1.5 mm wide, with a shallow channel on the upper side that
is broad at the tip and narrows regularly toward the base. The
lateral veins arise at a highly variable angle (40° to 65°) with the midrib,
and branch dichotomously at from slightly less than one half to nearly three
fourths of the distance from the midrib to the margins of the leaf.
This species seems to be clearly distinct from
all the others in the genus.
6.
Wenzelia melanesica Swing. Jour. Arnold. Arbor. 21:9. 1940.
Illus. Swing. loc. cit. pl. 2, figs. 6-8; pl. 3, figs.
1-3.
Type.—Solomon Islands,
Bougainville Island, Kugu-maru, Buin (Kajewski, No. 1907a).
Herb. Arnold Arbor., Harv. Univ.,
Cambridge.
Distribution.—Known only
from the type locality.
Small thornless tree
(up to 7 m); twigs glabrous, slender, 1-2.5 mm diam.; leaves thin, chartaceous,
glabrous, elliptical, with very numerous minute oil glands evenly distributed
over the whole surface, acuminate at the apex, which ends in a sharp point,
broadly and evenly rounded at base, 10-16 X 3.5-5.5 cm, margins entire or
slightly crenulate; lateral veins clearly marked below, 12-15 pairs, arising at
a large angle (70°-80°) with the midrib; petioles very short, 4-5 X 1-1.3 mm,
glabrous, not articulated with the blade, with a narrow, very shallow channel
on the upper side, the walls of which connect with the leaf margins; flowers
solitary or in small groups (2-3) in the axils of the leaves, pedicels with a
few sparse, short hairs, short (4-6 mm), with very small ciliate bracts at the
base, slender, expanding gradually into the funnel-shaped calyx; calyx 4-5
lobed, lobes broadly rounded or very bluntly pointed, 1.3-2 mm long, 1.5-2 mm
wide, and increasingly short-pubescent toward the margins, which are
abundantly short ciliate; corolla with 4-5 petals (immature in type specimen),
showing (in the bud) numerous oil glands evenly distributed; stamens 8-10;
pistil immature, ovary with 4-5 locules, each with 6 ovules; fruits 4-5 angled,
tapering to a sharp point at the apex, 4.5 cm long, 2.3 cm wide, peel showing
numerous, evenly distributed small (0.1-0.2 mm), slightly sunken oil glands;
seeds thin, flat or nearly so, ovate in outline, 9-13 X 6.5-9 X1.5-2 mm, dusky
gray, very thin at the edges, with a lighter colored, narrow, usually
subentire, marginal membrane; embryo monoembryonic.
This striking species of Wenzelia extends
the range of the subgenus Papualimo from New Guinea to the Solomon
Islands (Long. 156° E.). It resembles most closely W.
archboldiana from New Guinea but differs from it in having 4- to 5-angled
fruits tapering into a sharp point.
7. Wenzelia archboldiana Swing. Jour. Arnold Arbor.
21:7. 1940. Illus. Swing. loc. cit. pl. 2, figs.
1-5.
Type.—British New Guinea, lower
Fly River opposite Sturt Island in rain-forest (L. J. Brass, No. 8038,
fruiting branches). Herb. Arnold Arbor., Harv. Univ.,
Cambridge.
Distribution.—Southeastern
and eastern New Guinea. In addition to the type, two other
collections from northeastern New Guinea were seen: Morobe district, Ulap, 610
m alt. (Clemens, No. 6613, flowering branches), Herb. Natl. Arbor., Washington,
D.C.; and Morobe district, above Sattelberg (J. and M. S. Clemens, No. 1057,
branches with young fruit), Herb. Bot. Mus.,
Berlin-Dahlem.
Common name.—Archbold’s
wenzelia.
A thornless shrub or small tree, 3-5
m high; twigs glabrous, 2-4 mm diam.; leaves ovate-lanceolate or lanceolate,
16-22 X 7-9 cm, more or less abruptly acuminate at apex, cuneate at base,
margins entire or slightly crenulate, lateral veins 10-14 on each side, arising
at angles of 60°-70° with the midrib, usually branching dichtomously
[sic] at 1/2-3/5 of the distance to the margin, petioles very short, 4-8
X 1.5-2 mm (1/25-1/40 of the length of the leaf blade), with a shallow channel
on the upper side; flowers usually arising in pairs in the axil of the terminal
leaf, flower buds 2.3 X 3-4 mm, pedicels 4-5 X 0.8-1 mm, expanding gradually
into the funnel-shaped calyx 4-5 mm long and 3-4 mm wide at the top, calyx
lobes short (1 mm long, 3 mm wide), broadly rounded, minutely pubescent on the
back and margin; corolla white, petals 4 (or 5), measuring 15 X 4.5 mm in the
bud, dotted with small oil glands, glabrous, margins thin and slightly
scarious; pistil 12-12.5 mm long immediately after the petals fall, disk short,
0.9-1 mm high, 1.7 mm wide, abruptly constricted to 0.9-1 mm wide at junction
with ovary base; ovary clavate with a sterile narrowed base, 6-7 X 1.7-3 mm, 5-
(or 4-) locular, with 6 or 8 ovules in each locule, style cylindrical, 4.5-5 X
0.5-0.6 mm, not expanded at junction with ovary tip and only slightly expanded
at stigma junction, stigma cushion-shaped, 1 mm high, 2 mm wide; fruits
commonly paired in axil of uppermost leaf of fruiting twig, "lime green and
smelling of limes," depressed-globose, 2.5-3 X 3-3.5 cm, 5-locular (in type),
each locule bulged out (radial diam. 14-15 mm at junction with adjoining locule
and 17-18 mm midway between the two radial locule walls) making fruit
star-shaped in cross section with very blunt, rounded rays; seeds several (up to
6 or 8), crowded into each locule, flat and more or less curved, arranged more
or less radially with their long diameters parallel to the axis, 15-17 X 10-11
X 1.5-3.5 mm, brownish-gray (after soaking in formaldehyde solution),
monoembryonic.
The flowers of this species
were described from Clemens No. 6613, as the type specimen showed fruits
only. The most extraordinary character of the species—its numerous
flattened seeds—is one which it shares with three other species, W.
platysperma, W. paludosa, and W. melanesica. This
flattening of the seeds is undoubtedly not merely a result of the competition
for space by the numerous seeds that develop in each locule, but is an inherited
adaptation. These flat-seeded species of Wenzelia are
obviously related to Monanthocitrus cornuta, but it must be emphasized
that in Monanthocitrus the seeds are not merely very much flattened, but
are so compressed and crowded that they are distinctly saucer-shaped, concave
on one side and convex on the other, with a thin, more or less fimbriate
membrane almost entirely surrounding all but the attached side.
Furthermore, the seeds of Monanthocitrus are conspicuously spotted, a
character not found, so far as known, in any other plant belonging to the orange
subfamily.
The mature fruits of the type
specimen of W. archboldiana show a curious abnormality, having fusiform
cracks in the outer layer of the peel arising usually near the middle of the
dorsal wall of the locules just above the attachment to the
pedicel. The split in the peel reveals the more or less dried-up
oil glands and other tissues, but the inner peel apparently remains
intact. Sometimes these fusiform cracks occur between the locules
of the fruit, although usually not so abundantly.
The depressed-globose, ribbed, green fruits
(said to smelt like limes) usually appear in pairs in the axil of the uppermost
leaf on the fruiting twig—in striking contrast to W. platysperma, where
the more or less elongated subglobose or ovate pink fruits are borne singly,
and hang down apparently at the tip of the twig, although in reality in the
axil of the uppermost leaf.
This species has
been named in honor of Mr. Richard Archbold, who organized and led three great
collecting and exploring expeditions to New Guinea. He employed
expert collectors, among them L. J. Brass, with the result that many herbarium
specimens of the remarkably interesting plants of New Guinea were brought
back.
SPECIES INADEQUATELY KNOWN AND OF UNCERTAIN RELATIONSHIPS
8. Wenzelia dolichophylla
(Lauterb. & K. Schum.) Tan. Jour. Arnold Arbor. 9:139. 1928.
Citrus dolichophylla Lauterb. & K. Schum. in Schum. & Lauterb.
Fl. Deut. Schutzgeb. 377. 1901.
Type.—Northeastern New Guinea, tributary river No. 9, of Ramu River (Lat.
4° 20' S., Long. 144° 38' E.), high forest, altitude 60 m (Lauterbach, No.
3108). Herb. Bot. Mus.,
Berlin-Dahlem.
Distribution.—Known also
from the upper course of the Gogol River, northeastern New Guinea, and a
sterile specimen (doubtfully this species) reported by Lauterbach from Etna Bay
(Lat. 4° S., Long. 134° 30’ E.) in northwestern Dutch New Guinea (now West
Irian, Indonesia).
The original description,
translated, reads as follows: "Shrub with numerous slender young fruiting
branches, glabrous, terete; petioles very short, furrowed above; leaves
ovate-lanceolate or ovate-oblong, very acutely attenuate-acuminate, glabrous
on both sides, coriaceous; immature fruits long-pedicellate, subglobose,
somewhat acute, glabrous. A shrub 2 m high. The
branches are 20-25 cm long and 2-3 mm thick at the base; they have gray to brown
bark. The petiole is 3-5 mm long, no articulation of the leaf blade
with the petiole is visible; leaves 15-25 cm long and at the middle or lower
third 5.5-8.5 cm broad, with 14-18 pairs of veins arising at right angles from
the mid-rib, less prominent above than below; gray above, yellowish below, dull
on both sides. The pedicel is 2-2.5 cm long. The
immature fruits, still green, have a lemon-like odor."
Lauterbach reported (1918, p. 262) on a specimen
from the upper course of the Gogol River (Lauterbach, No. 1088) collected in
fruit, November 26, 1890, as follows: "The nearly ripe fruits are red,
lemon-shaped, 5.5 cm long and 3.5 in diameter." These are the
largest fruits yet reported in this genus and the red color is also
distinctive. Swingle examined the type specimen in the herbarium at
Berlin-Dahlem many years ago and found no trace of pulp-vesicles, the locules
of the dry fruits being filled with a brown, brittle, resin-like substance.
9. Wenzelia grandiflora
(Lauterb.) Swing. Jour. Wash. Acad. Sci. 28:532. 1938. Citrus
grandiflora Lauterb. Nova Guinea 8:293. 1910; Monanthocitrus
grandiflora (Lauterb.) Tan. Jour. Arnold Arbor. 9:139.
1928.
Type.—Central-western New Guinea
[West Irian, Indonesia], swampy primeval forest, Bivak Island, in Noord Fluss
[Lorentz River] (Lat. 5° 0' 25" S., Long. 138° 39' E.), (Versteeg, No.
1788). Herb., Utrecht.
Distribution.—Known only from the type locality.
The original description of Citrus
grandiflora by Lauterbach reads, translated, as follows: "Shrub 1-2 m;
branches slender, spiny, 2-4 mm thick, terete, flattened when young; spines 2,
about 1 cm long, borne in the axils of the leaves; leaves lanceolate,
alternate, acuminate [at the tip], acumen acute, broadly pointed at base,
15-20 cm long, 5-7.5 cm wide, coriaceous, margin crenulate-subdentate, veins
more prominent below, 12-13 pairs, ascending, anastomosing; petioles 5 mm long,
subterete; flowers solitary (?), axillary (?), with short pedicels; calyx
cupulate, 4 mm long, 5-merous, lobes triangular, subacute or somewhat truncate,
2 mm long; petals 5, white, lanceolate, broadly pointed ('late acutis') [at the
tip], attenuate at base, 35 mm long, 11 mm wide; stamens 8, free, filaments 11
mm long, enlarged at the middle, anthers oblong, 3.5 mm long; disk 1.5 mm long,
somewhat pentagonal; ovary substipitate, ovate, 3 mm thick, 5-locular, locules
with many ovules; style 1 mm thick, 6 mm long, stigma dilated, truncate,
pentagonal."
Unfortunately, the fruits of this
species are unknown, but in its flower, leaf, and petiole characters it agrees
very well with Wenzelia. It is the only species of
Wenzelia that has paired spines. However, the number of
spines is variable in the genus, six species having no spines and two, W.
paludosa and W. dolichophylla, having solitary spines, although the
latter species is sometimes spineless.
Wenzelia grandiflora was first described as a species of
Citrus. Many years later it was transferred to the monotypic
genus Monanthocitrus because the two showed agreement in having paired
spines in the axils of the leaves. However, the flowers of
Monanthocitrus are very much smaller than those of any species of
Wenzelia, the petals being only one seventh as long as those of W.
grandiflora! Until fruits with mature seeds are found this
latter species must stay in the genus Wenzelia.
VII. Monanthocitrus Tan.
VII. Monanthocitrus Tan. Jour. Arnold Arbor. 9:138. 1928.
Illus. fig. 3-11.
Type
species.—Citrus cornuta Lauterb. = Monanthocitrus cornuta
(Lauterb.) Tan.
Distribution.—Southwestern Dutch New Guinea [West Irian, Indonesia]: near
Alkmaar (Lat. 4° 40' S., Long. 138° 44' E.) on the Lorentz River and Signal
Hill, a few miles west of Alkmaar (see van Nouhuys, 1923, pl.
4).
Common name.—Spotseed-lime (or
monanthocitrus).
Twigs slender, slightly
angled when young, soon cylindrical, internodes long (2-4 times as long as the
petioles); spines paired or single (rarely 3, or wanting), very slender, about
half again longer than the petioles; leaves simple, 14-22 cm long, elliptical,
caudate-acuminate at the tips, broadly cuneate at the base, glabrous, margins
entire or slightly wavy; lateral veins 10-15 pairs, visible on both sides;
petioles very short (shorter than the spines and less than 1/20 as long as the
leaf blade), wingless, flattened on the upper side, not articulated with the
leaf blade; flower buds ovoid or ellipsoid, borne on slender pedicels as long or
longer than the petioles; flowers small; calyx cupulate, 5-lobed, lobes acute,
short; petals 5, shorter than the petioles; ovary globose, very small, with 5
locules, each locule with 6 (or sometimes 8?) ovules; style thick, twice as
long as the ovary, stigma 5-lobulate; fruits globose or slightly ovoid, with
thin peel showing numerous slightly depressed oil glands; locules 5, lined with
smooth, cartilaginous walls, without pulp-vesicles, but containing 6 (or
sometimes 8?) seeds surrounded by mucilaginous fluid; seeds packed tightly in
the locule, much flattened, often concave (like a shallow saucer), testa with
many small irregular dark red-brown spots; a very thin irregularly lobed
membrane borders the seeds except on the attached
side.
Monanthocitrus has leaves very
much like those of Wenzelia dolichophylla and several other species of
that genus. Monanthocitrus is most closely related to the
species of Wenzelia belonging to the subgenus Papualimo, which
have flattened seeds bearing on the unattached sides very thin, more or less
torn membranous margins. However, no species of Wenzelia has
the much-spotted seeds (unique in the orange subfamily) found in
Monanthocitrus, which also has flowers very much smaller than those of
any species of Wenzelia—with petals only one-fifth to one-seventh as
long! Doubtless Monanthocitrus arose from an ancestral form
much like that of some of the species of Wenzelia of the subgenus
Papualimo, but it has traveled so far on the evolutionary highway that it
apparently constitutes a good genus today. The leaves are also
much like those of Clymenia polyandra (a True Citrus Fruit Tree of the
subtribe Citrinae), which has petioles of about the same length in relation to
the leaf blade as those in Monanthocitrus, i.e., about one-twentieth as
long.
Thanks to the kindness of Dr. H. J. Lam,
director of the Rijks Herbarium at Leiden, Holland, Swingle was able to study
the type specimen on which Citrus cornuta Lauterb. was based, upon which
species Tanaka based his genus Monanthocitrus. This type
specimen was collected by Versteeg (No. 1551) at Alkmaar in New Guinea in
August, 1907. It carries the fruit described by Lauterbach, showing
curious slender, needle-like outgrowths from the tip of each carpel.
The fruit was still intact and when opened it showed no trace of
pulp-vesicles.
Monanthocitrus cornuta
(Lauterb.) Tan. Jour. Arnold Arbor. 9:138. 1928. Citrus cornuta
Lauterb. Nova Guinea 8:292. 1910. Illus. fig. 3-11.
Type.—Southwestern Dutch New Guinea [West Irian, Indonesia], near Alkmaar
(Lat. 4° 40' S., Long. 138° 44' E.) on the Lorentz River (Versteeg, No. 1551),
Rijks Herb., Leiden.
Distribution.—Known only from the type locality and from Signal Hill, a few
miles west of Alkmaar.
Lauterbach's
description of Citrus cornuta (with slight changes in the order of
arrangement) reads, translated, as follows: "Shrub about 1 m high; branches
slender, flower-bearing ones 2-4 mm thick, terete, spiny; spines 2 or 3, about
1 cm long, in the axils of the leaves; leaves ovate-oblong or lanceolate,
attenuate-acuminate, acumen up to 2 cm long, obtuse, sometimes emarginate,
coriaceous, cuneate at base, entire or wavy margined, veins more prominent
below, lateral veins in 10-12 pairs, ascending, thickly anastomosing by arches
near the margin; petioles semiterete, scarcely channeled above, 5-6 mm long;
flowers solitary, axillary, pedicellate; pedicels 5-8 mm long, subtended by 2-3
acute bracts, 1 mm long; calyx cupulate, 5-merous, lobes acute, 1 mm long,
slightly ciliate under the lens; petals 5, ovate, subacute, about 5 mm long,
whitish flesh-colored ('albis carnosulis'); stamens 10, free, filaments 1.5 mm
long, anthers oblong, 2 mm long; disk annular; ovary globose, 1 mm diam.,
5-locular, 2 ovules [? see below] in each locule; style thick, 2 mm long, stigma
5-lobed; fruit globose, rose-colored, 2.5 cm diam. when dry, fleshy, the apex
with 5 solitary mammillae, each crowned by a single spine [-like appendage] up
to 2 cm long.…
"The long spines that form what
may be called horns are very peculiar; as far as can be determined from dry
material they arise from wart-like protuberances at the apex of the fruit and,
in the dry state, fall off easily; 3 are intact; only the points of attachment
of the other two are seen. No trace of their origin can be seen on
the ovaries."
Swingle examined the type
specimen of this species (Versteeg, No. 1551, collected at Alkmaar, New Guinea,
August 2, 1907), lent to him by Dr. H. J. Lam, director of the Rijks Herbarium
at Leiden. It consists of a branch about 20 to 25 cm long, with six
leaves (see fig. 3-11). It bears one fruit, 25 by 32 mm, with
well-marked oil glands. When described by Lauterbach in 1910 the
specimen still had three needle-like appendages arising from the top of the
fruit and the bases of two more. Doubtless there were originally
five of them, one arising from each locule of the fruit. These
needle-like appendages are 20 to 22 mm long and very slender, about 0.6 to 0.75
mm in diameter at the base and 0.3 to 0.4 mm at the tip (measured when dry);
they have about 20 to 22 shallow furrows, with narrow ridges between, extending
lengthwise. They arise at the apex of the fruit from wrinkled,
glabrous elevations (broadly conical) of the peel about 2.5 to 3.5 mm wide and
2 to 2.5 mm high; one of these bases is found on each segment.
These remarkable needle-like outgrowths of the locules of the fruit look like
slender projections of the peel and have the same color as the peel, but do not
show oil glands. The largest leaf blade measures 20.5 by 7 cm; two
smaller leaves are 16 by 5.5 cm and 15 by 5 cm. The petioles are
channeled on the upper side, 7 to 8 by 1.5 to 1.6 mm, and not articulated with
the leaf blade; the paired spines are straight, 3 to 12 by 0.5 to 0.8
mm. The single fruit is oval, 30 mm long and 25 mm wide when dry;
when swelled by soaking in hot water it measured 32 mm long and 28 to 30 mm
wide. The greatest length from the base to the tip of one of the
five conical bases on which the needle-like outgrowths arise is 34
mm. The dried fruit is buff to cinnamon in color (see Ridgway,
1912, pl. 39). The pedicel is 10 mm long with two very small bracts
attached 2 mm above the base. The calyx lobes are reflexed when
dry, but on soaking they extended out nearly at right angles to the pedicel,
making the salverform calyx 6 to 7 mm wide and about 2.5 mm high.
The calyx lobes are about 3 mm long, thick and bluntly rounded at the tips, and
become very thin at the extreme tips and sparingly ciliate. When
the fruit was cut open, the peel was seen to be thin, only 1 to 1.5 mm including
the locule wall. The outer peel is 0.6 to 0.8 mm thick; the locule
walls are stiff, cartilaginous, and smooth inside. The seeds, six
(or sometimes eight) in each locule, are crowded into a bundle nearly filling
the locule, which also contains mucilaginous gum, but no
pulp-vesicles! The seeds are very thin, often curved (like a
shallow saucer), and have thin fimbriate margins, cream, buff, or ivory-yellow
in color on all but the attached side of the seed. Including this
membrane, the seeds measure 10 to 11 by 7 to 8 by 1.5 to 2 mm. The
seed proper measures about 8 to 9.5 by 7 to 8.5 mm and is covered with an
ivory-yellow testa which becomes fawn to army brown (Ridgway, 1912, pl. 40)
when wet. This testa shows many (10 to 20 or more) angular, dark
red-brown spots, varying greatly in size from 0.5 to 2.5 mm, usually narrow if
long. On well-soaked seeds these spots varied from dark vinaceous to
Indian red (Ridgway, 1912, pl. 27). The inside layer of the testa
in contact with the cotyledons of the embryo shows numerous very small, curved,
vermicular markings of vinaceous brown (Ridgway, 1912, pl. 39). The
seeds contain a single very thin embryo, oval in outline, 8 to 9 by 7 to 8 mm,
and 1 to 1.5 mm thick, vinaceous fawn to fawn color (Ridgway, 1912, pl.
40). No oil glands can be detected in the cotyledons by superficial
examination. The very small radicle projects out 0.5 to 0.8 mm at
the base of the cotyledons. The attached edge of a well-soaked seed
showed a more or less rugose ariloid tissue, 6 to 7 mm long and about 2 mm wide,
which projects 1.5 to 2 mm beyond the seed.
A
paratype specimen of M. cornuta (Versteeg, No. 1551), also collected at
Alkmaar, New Guinea, on August 2, 1907, preserved in the herbarium of the
Botanical Museum at Berlin-Dahlem, was photographed by Tanaka, who kindly sent
Swingle a print. It is a single twig with one flower bud, 4.5 by 3
mm, on a slender pedicel about 7 mm long. One leaf measures 2l by 7
cm; another, lacking the acuminate tip, measures 18.5 by 8.2 cm, with a petiole
8 by 1.5 mm. The spines are all paired, straight, very slender, 8
to 14 mm long and 0.6 to 0.8 mm at the base and tapering to a very acute
point. The lateral veins arise at a large angle with the midrib,
about 65° to 75°, in the middle part of the leaf blade.
The remarkable needle-like protuberances that
arise at the tips of each carpel of the fruit of the type specimen are still
imperfectly known.4 In a supplementary
note on this plant Lauterbach stated (1912, p. 825) that a specimen collected
by von Roemer (No. 633) at Signal Hill, a few miles west of the type locality
at Alkmaar, "had fruits with no trace of the peculiar spines. Very
likely it was a question, in case of the original specimen of Versteeg, No.
1551, of a mere malformation that was nevertheless of remarkable regularity."
VIII. Oxanthera
Montrouzier
VIII. Oxanthera
Montrouzier, Mém. Acad. Sci. Lyon 10:186.
1860.
Type species.—Oxanthera
fragrans Montr.
Distribution.—New
Caledonia and Art Island (80 km to the northwest of New
Caledonia).
Common name.—False orange
(or oxanthera).
Thornless small trees or
shrubs with thick but small 1-foliolate or simple leaves (3-10 cm long),
usually becoming glaucous; petioles either wingless or narrowly winged,
articulated with the leaf blade in 3 species, not articulated in the other
species; flower borne singly or in 1-3-flowered racemes in the axils of the
leaves; calyx and corolla 5-merous; stamens 3-4 times as numerous as the
petals; ovary with 2-7 locules, each locule containing several ovules, usually
6; fruits ellipsoid, small, about 2.5 X 1.5
cm.
Oxanthera is a very distinct genus
of Minor Citroid Fruit Trees, subtribe Triphasiinae, and differs strikingly
from all the other genera of this subtribe in having glabrous, glaucous leaves
bluntly rounded or retuse at the tip and cuneate at the base, borne on
spineless twigs. The leaves are thick and coriaceous in all the
species except one. To judge from the thick leaves that become
glaucous as they develop, the plants of this genus are very probably somewhat
xerophytic, like Microcitrus of Australia, but are not so well adapted to
withstand long-continued drought and hot, dry winds as the truly xerophytic
Eremocitrus, also of Australia.
All the
species of Oxanthera agree in having large, orange-like flowers, and
glabrous, more or less glaucous leaves, which usually are rather blunt at the
tip and cuneate at the base. All species are thornless, and
apparently all have more or less elongate fruits that are longitudinally ribbed
at least when young. Probably no other genus except Poncirus
in the whole subfamily is so easily recognized at first glance.
In spite of this fact, this genus, containing four species (see key) shows unusual range of variation in other characters that in
many groups of plants are extremely constant and have high taxonomic
value. Three of the four
species of Oxanthera have unifoliolate leaves with clearly articulated
petioles that are usually wingless, but are plainly but narrowly winged in one
species. One of the species has a hypomerous ovary with only two
locules, another has an isomerous ovary with five or six locules, whereas a
third species has a hypermerous ovary with seven locules. Better
material of O. aurantium is urgently needed to learn whether the ovary
is always 2-locular; if so, it might well be made the type of a
subgenus. For the present it is left in the genus as an anomalous
species that, nevertheless, is clearly related to the other three species.
It is probable that Oxanthera is a highly
specialized xerophytic genus that has developed from the common ancestor of the
genera Wenzelia and Monanthocitrus, both of which share with
Oxanthera the character of having usually six ovules in each locule of
the ovary. Oxanthera has acquired a distinctly xerophytic
facies but, unlike the extremely xerophytic Eremocitrus, one of the True
Citrus Fruit Trees of Australia, which has very small flowers, Oxanthera
has large, white, fragrant flowers very like those of Citrus.
1. Oxanthera fragrans Montr.
Mém. Acad. Sci. Lyon 10:186. 1860. Citrus oxanthera Beauv. Ann.
Soc. Bot. Lyon 26:14. pro parte.
1901.
Type.—L'ile Art (about 80 km
northwest of the north end of New Caledonia) (Montrouzier). Herb.
Faculte de Méd. et Pharm. de Lyon.
Distribution.—Art Island, New
Caledonia.
Common name.—Fragrant
oxanthera.
The original description by
Montrouzier, translated, reads as follows: "Shrub 6-7 ft. [1.8-2.13 m] high,
beautiful, twigs terete, gray. Leaves obovate, apex acuminate,
subcrenulate, thick, very much dotted [with oil glands], 2-2.5 in. [5-6.3 cm]
long (including the petiole), 6-7 lin. [1.2-1.4 cm] wide. Peduncles
axillary, 1-flowered, shorter than the leaves. Flowers white, the
size of those of Citrus Aurantium, and very similar to them, very much
dotted [with oil glands], exhaling a penetrating odor. Fruit
elongated. Is this Limonia lucida Forst.? This
beautiful shrub flowers in November. It is found in the
forest. The natives call it Dongan na diet, 'Orange Tree of
the Forest.' "
As the genus Oxanthera
was originally monotypic, being founded on O. fragrans alone,
Montrouzier's description of the genus applies to the species as
well. It reads, translated, as follows: "Flowers 5-merous, with
imbricate aestivation, stamens 16-20, free. Anthers erect,
bilocular, incurved, mucronate at the apex. Fruit with 5-6 locules,
with many seeds in each locule. Style elongate, thick,
cylindric. Stigma obtuse. Leaves 1-foliolate,
articulated with the tip of the petiole, petioles wingless. A
spineless shrub."
Tanaka in discussing O.
aurantium (1928b, p. 44) stated: O. fragrans has petals
clearly marked with translucent points, a glabrous calyx, oblong anthers with
only a small mucron at the apex, an oblong-oval ovary much furrowed ['très
creusé’], and a very short style." It differs in all these points
from O. aurantium, with which Beauvisage confused it, since he included
descriptions of material of both species in his Citrus oxanthera.
The type specimen photographed by Tanaka shows petioles 4.5 to 7 mm long,
articulated with the leaf blade, entirely wingless but apparently with a
shallow channel on the upper side.
This plant
is said to be a beautiful fragrant shrub and may perhaps prove of value as an
ornamental flowering shrub for subtropical climates. To judge from
its foliage, it is probably somewhat drouth-resistant [sic].
The petals, with densely crowded oil glands, may possibly yield an essential oil
for perfume.
2. Oxanthera
aurantium Tan. Bul. Soc. Bot. France 75:712. 1928. Citrus
oxanthera Beauv. Ann. Soc. Bot. Lyon 26:14. pro parte. 1901.
Illus. Guillaumin, Les Citrus Cult. Sauv. 60.
1917.
Type.—New Caledonia, Gomonen near
Gatope on dry hills (Vieillard, No. 2378). Herb. Mus. Hist. Nat.,
Paris.
Distribution.—Known only from
the original collections.
Common
name.—Orange flower oxanthera.
Tanaka’s
original description reads, in translation, as follows: "A shrub, twigs terete,
very leafy, young twigs pubescent; leaves small, oblanceolate, not glandular [?,
‘englandulosis’], chartaceous, channeled, with many veins, lateral veins
curved, arising from the midrib at more or less wide angles; apex obtuse or
often emarginate, base attenuated, short-petiolate; petioles cylindric, not
articulated [with the leaf blade]; inflorescences pubescent, 1-3-flowered
racemes; flowers large, equaling Citrus in size, petals without oil
glands, minutely papillose; calyx scutellate, pubescent without, lobes rounded,
inflexed; stamens free or in part cohering, filaments much longer than the
style, canaliculate, minutely papillose on the outside, anthers rather large,
cordate-acuminate; ovary ovoid-attenuate, glabrous, style equaling the ovary
[in length], straight, cylindric, rather thick, stigma ellipsoid, disk annular,
short, stigma ellipsoid, broader than the style."
Swingle examined two cotypes from the original
type collection of this species (Vieillard, No. 2378), one in the herbarium of
the Museum of Natural History, Paris, and the other, an excellent flowering
specimen, in Gray Herbarium, Harvard University, Cambridge,
Massachusetts. The latter specimen shows completely thornless
twigs, pubescent when young but losing the hairs and appearing glaucous with
grey wax when older; leaves simple, thick, coriaceous, oblanceolate, 2-4 X
0.5-1 cm, apex rounded, usually rather deeply emarginate, margins minutely
crenulate-dentate above the middle, upper surface dotted with minute oil
glands, about 0.5 mm apart, which are not translucent on dry specimens, midrib
slightly raised on the upper surface, strongly salient below, slightly
pubescent near the base of the leaf, lateral veins visible only on the under
surface, curved and more or less swollen and salient, arising sometimes at an
angle of 35°-45°, sometimes at an angle of 75°-80°, with the midrib, soon
curved backward toward the base of the leaf and anastomosing especially in the
upper part of the leaf; petioles pubescent, not articulated with the leaf
blade but merging imperceptibly into it, flattened above and showing two raised
lines that continue the leaf margins, running almost to the base of the
petiole, but showing no green color for 2-3 mm from the base; peduncles at
flowering densely covered with gray hairs, with several leaf-like bracts,
making the peduncle appear like a short leafy twig; pedicel very short (3-4
mm), not clearly demarcated from the calyx, which is densely gray-pubescent
without; flowers like Citrus except for the gray-pubescent calyx and
minutely papillose petals.
Tanaka
(1928b, p. 712) noted that this species differs greatly from O.
fragrans in its pubescent new growth, narrower leaves with many lateral
veins arising at a wide angle with the midrib, and entirely different flower
parts. Beauvisage (1901, pp. 11, 12) studied several specimens of
Vieillard's No. 2378 (the type collection on which O. aurantium is based)
and his report, translated, reads as follows: "I dissected several rather badly
preserved flowers and found 14 to 16 stamens with mucronate anthers and connate
filaments except in a very young bud, where the filaments, still short, were
free. Similarly I noted in the ovaries a few differences which
could be due to the more or less advanced stage of development; the ovary was
ovoid or conical; the style articulated [with the ovary] or not; the stigma
channeled-ellipsoid or not; the locules of the ovary, usually with several
ovules ('pluriovalées'), seem sometimes to be biovulate."
A good specimen of the type collection
(Vieillard's No. 2378) in the Gray Herbarium at Harvard University provided a
flower which Swingle examined. It was restored by prolonged soaking,
then imbedded, cut into serial sections, stained and mounted permanently by a
modification of Juel's method (see Swingle, 1939b, p. 270).
The ovary was skillfully cut by Swingle's assistant in this part of the work,
Albert H. Tillson, so as to show half the ovary cut into many serial
longitudinal sections and the remaining (longitudinal) half cut into still more
numerous cross sections. These sections showed clearly that the
ovary was cylindric, about 3.5 mm long after the dehiscence of the style, and
0.9 to 1.2 mm in diameter (with two shallow furrows in the plane of junction of
the carpels and two low ridges on the dorsal walls), narrowed abruptly at the
base and fitting rather closely into a conical cavity in the top of the disk; it
contained two long, slender locules about 2.4 by 0.3 by 0.4 mm. The
ovary was narrowed gradually at the top into the style, 0.5 to 0.6 mm in
diameter at the point of dehiscence. The serial cross sections
showed altogether six ovules in the locule, arranged in two ranks, along with
abundant trichomes arising from the flattened placenta that ran from bottom to
top of the locule cavity along its dorsal wall.
This species differs widely from its congeners
in having a hypomerous ovary with only two locules. Of the other
species, one is isomerous with five or six locules and another is hypomerous
with seven locules. O. aurantium also differs radically from
its congeners in having simple instead of unifoliolate leaves. Its
relationships are discussed above.
This species, collected
by Vieillard on arid hills in New Caledonia, shows signs of being a rather
pronounced xerophyte able to grow in dry situations. The thick
leathery leaves remind one of the leaves of Eremocitrus grown on
well-watered soil but are not so gray-green in color and show a different
structure on the upper and lower surface, a difference that is almost lost in
Eremocitrus.
3.
Oxanthera undulata (Guill.) Swing. Jour. Wash. Acad. Sci. 30:83.
1940. Citrus undulata Guill. Bul. Soc. Bot. France 85:304.
1938.
Type.—New Caledonia, upper Dothio
Valley (Balansa, No. 3054). Herb. Mus. Hist. Nat.,
Paris.
Distribution.—Known only from
the type locality in northern New
Caledonia.
Common name.—Wavy-leaf
oxanthera.
The original description of this
species reads, translated, as follows: "Tree 5-6 m high, very glabrous; leaves
thin, ovate-lanceolate, up to 9 X 4 cm, cuneate at the apex and base, with
undulate margins, nerves very slender, almost immersed, petiole articulated
[with the lamina], slender, 5-10 mm long, [wingless and] not at all margined;
flowers not seen; fruits ovate, 2.5 X 2 cm, embryo with white cotyledons…"
The notes which follow the Latin diagnosis read:
"Excessively rare; a species well characterized, although imperfectly known, by
its thin leaves with strongly undulate margins. The fruit resembles
that of C. neo-caledonica in form." Unfortunately the number
of locules in the fruit is not stated.
To
judge from its large thin leaves, this species may perhaps prove to be a
mesophyte rather than a xerophyte, as are the other 3 species in
Oxanthera.
4. Oxanthera
neo-caledonica (Guill.) Tan. Bul. Soc. Bot. France 75:713. 1928.
Citrus neo-caledonica Guill. in Lecomte, Not. Syst. 2:130
(diagnosis). 1911; and ibid. 2:390 and 406 (Latin name).5 1913. Illus.
Guillaumin, Les Citrus Cult. Sauv. 61. 1917; fig. 3-12 this work.
Type.—New Caledonia, Poume Peninsula (Balansa, No. 3374). Herb.
Mus. Hist. Nat., Paris. Cotype: Same collection (Balansa, No.
3374). Herb. Bur. Sci., Manila.
Distribution.—Known only from the type locality (Lat. 20° 30' S., Long. 164°
8' E.).
Common name.—False orange (or
large-leaf oxanthera).
Guillaumin's detailed
description of this plant reads, in translation, as follows: "Spineless shrub, 1
m high, with whitish flowers; young branches with a waxy bloom that gradually
disappears. Leaves very coriaceous, glabrous even when very young,
lanceolate or obovate, 6-8 cm long (including the petiole), 1-4 cm wide,
narrowed toward the base, attenuate toward the apex, but not acute when
lanceolate, suborbicular or orbicular when obovate, sometimes
emarginate. Midrib and lateral veins prominent on both sides but
especially on the underside (like veins of the hand). Petiole
distinctly articulated [with the leaf blade], 1-2 cm long, slightly winged (less
than 4 mm). Flower [bud] (a single one) terminal on a young branch,
spherical (14 mm diam.); pedicel 7 mm long, cylindric, glabrous; sepals 5, 2 mm
long, nearly round; petals 6, unequal, ovate (12 X 6-12 mm), thick and fleshy,
with inconspicuous, small, pellucid oil glands; stamens 24, unequal (8-11 mm),
[arranged] very irregularly in fascicles, filaments rather fleshy, not
flattened, anthers triangular-elongate, 2.5 mm long, the locules diverging
slightly at the base; ovary subcylindric (3.5 X 3 mm), 7-ribbed, with 7
segments; style cylindric, 2 mm long, stigma cylindric. Fruit
(immature) ovoid-subcylindric, supported on pedicels covered with whitish bloom,
more than 1.5 cm long and 2 mm diam.; more mature fruits 2.5 X 1.5 cm, with 7
segments, peel thin, pulp mucilaginous, seeds of unknown color.
"Presqu'ile de Poume, in volcanic soil, May, 1871
(Balansa, No. 3374).
"Although the color of
the embryo is not known, the affinities [or this plant] seem to be especially
with Citrus Aurantium L. but the form and the aspect of the leaves, of
the flowers and of the ovary, clearly distinguish this species."
Tanaka (1928b, p. 44) noted that the
flowers are like those of O. fragrans and O. aurantium but that
"the surface of the petals is completely uniform [not papillose or marked with
evident oil glands], the ovary is shorter, the stigma not dilated and the disk
much larger." He went on to say: "The fruit is oblong, sometimes
curved, pointed, with superficial ribs, with about 7 segments, containing large
seeds with a single very hard embryo. The pulp-vesicles are not
very much developed and are gummy." He was of the opinion that the
lack of true pulp-vesicles excludes this species from the True Citrus Fruit
Trees even though it does have more than twice as many stamens as petals.
This species, as Tanaka remarked, shows a higher
organization than most of the other Minor Citroid Fruit Trees. It
has seven locules in the ovary and, in common with the other species of
Oxanthera, more than twice as many stamens as petals, instead of only
twice as many, as is characteristic of the other genera of the subtribe
Triphasiinae.
IX. Merope
Roem.
IX. Merope Roem. Syn.
Hesper. 1:44. 1846.
Type species
(Monotype).—Citrus angulata Willd. = Merope angulata (Willd.)
Swing.
Distribution.—Java, Burma; also
New Guinea (fide Tanaka, 1931b, p. 10).
Small tree, twigs with stout spines paired or
single; leaves simple, thick, coriaceous; petioles short, wingless; flowers
solitary, axillary; calyx cupulate, 5-lobed; petals 5, imbricate,
oblong-lanceolate, acute; stamens 10, free, anthers linear, as long or longer
than the filaments; ovary ovoid, 3-locular, ovules 4 in each locule; fruit
triangular in cross section, ovoid-acuminate, with very long, flattened seeds
narrowed at the upper end to a blunt point, borne 1 in a segment and immersed
in a glutinous, mucilaginous fluid.
This
remarkable genus is found widely scattered over the East Indies in brackish
swamps. It is outstanding because of its buckwheat-shaped fruit,
containing very long seeds. It is a highly specialized genus of the
subtribe Triphasiinae, related to Pamburus and Paramignya, but
not closely.
Merope angulata (Willd.)
Swing. Jour. Wash. Acad. Sci. 5:420. 1915. Citrus angulata Willd.
Sp. Pl. ed. 4. 3(2):1426. 1801; Limonellus angulosus Rumph. Herb.
Amboin. 2:110. 1741; Sclerostylis spinosa Blume, 1825;
Glycosmis spinosa (Bl.) Dietr. 1840; Merope spinosa (Bl.) Roem.
1846; Limonia angulosa Wt. & Arn. ex Miq. 1859; Atalantia
longispina Kurz, 1872; Gonocitrus angulatus (Willd.) Kurz, 1874;
Paramignya angulata (Willd.) Kurz, 1875; Atalantia spinosa (Bl.)
Koorders (non Hook. f.), 1912; A. angulata (Willd.) Engl. Die Nat.
Pflanzenfam. 19a:330. 1931. Illus. Rumphius, loc. cit. pl.
32; Kurz, Jour. Asiat. Soc. Bengal 43(2):pl. 18. 1873; Valeton, Icon.
Bogor. pl. 348. 1912; Swingle, loc. cit. figs. 1-2; idem, in
Bailey, Stand. Cycl. Hort. 4:2038, fig. 2362. 1916; idem, in
Engler, loc. cit. 329, fig. 151; fig. 3-13 this work.
Type.—None found; based on Rumphius' description and figure cited
above.
Distribution.—Java, Burma, in
tidal swamps; also New Guinea (fide Tanaka, 1931b, p.
10).
Common name.—Merope (or
kigerukkan, so-called at Noessa Kambangan Island, Java).
Leaves coriaceous, inconspicuously veined,
alternate; petioles simple. Twigs with very strong spines, often in
pairs. Flowers borne singly or in pairs (rarely in few-flowered
clusters) in the axils of the leaves; ovary stalked on a rather tall disk,
3-5-celled, with 4 pendulous ovules in each cell; stamens 10, free, anthers
linear-oblong. Fruits strongly angled, triangular in cross section;
cells filled with a sticky, mucilaginous fluid (without true pulp-vesicles);
seeds very large, flattened, reniform, caudate at the tip where
attached. Cotyledons in germination aerial, not increasing in
size; first foliage leaves alternate, broadly ovate. A large shrub
or small tree (not a climbing shrub), growing on the seashore in tidal forests
or mangrove swamps from the mouth of the Ganges to the Moluccas.
The seeds, often 2 in each of the flattened locules of the fruit, are about 35
X 15 X 5 mm, largest known in the orange subfamily.
This
remarkable plant, the sole species of the genus Merope,differs
strikingly from all other species of the orange subfamily except Severinia
lauterbachii in having fruits triangular in cross section, with long flat
seeds tapering to a blunt point. It is a very spiny salt-loving
shrub or small tree found in tidal marshes in Java and Burma, and in the East
Indian Archipelago. As is usual with halophytes (plants able to
live in more or less salty water), M. angulata has thick and leathery
leaves. Buds of the
kigerukkan, inserted on grapefruit, grew vigorously in the greenhouse of
the former Bureau of Plant Industry at Washington, D.C., and soon flowered (see
fig.
3-13), but finally showed signs of a bad bud union, became stunted, and
died.
GROUP B. THE TRIPHASIA
GROUP
This group contains two genera with a total of
four species, all of them shrubs or small trees with small ovoid or globose
fruits, bright-colored (red) in Triphasia and orange-like with a thick
peel dotted with oil glands in Pamburus, but always without pulp-vesicles
in both genera. The ovaries are 3- to 5-locular, and each locule
contains one or two ovules. The leaves are simple in two of the
species of Triphasia, as well as in Pamburus, and trifoliolate in
the other species of Triphasia; they are rather thick, more or less
glaucous, and inconspicuously veined. Little is known as yet about
the relationships of these two genera with the other genera in the subtribe
Triphasiinae.
X. Triphasia Lour.
X. Triphasia Lour. Fl. Cochinch. 152. 1790. Echinocitrus
Tan. Jour. Arnold Arbor. 9:137.
1928.
Type species.—Triphasia
aurantiola Lour. = T. trifolia (Burm. f.) P.
Wils.
Distribution.—Southeastern Asia,
East Indies, Philippines, New Guinea; widely cultivated in tropical and
subtropical regions.
Common
name.—Limeberry (or triphasia).
Shrubs or
very small trees, 1-2 m high, with paired axillary spines; leaves usually very
small (except in T. grandifolia), simple or 3-foliolate, rather thin and
without evident reticulation, numerous oil glands visible on both faces;
petioles very short (1/10-1/20 the length of the leaves), wingless, pubescent,
flattened and slightly channeled on the upper surface, not articulated with the
leaf blade; flowers single, or in groups of 2 or 3 in the axils of the leaves;
pedicels slender; calyx 3-5-merous, cupulate, persistent; corolla 3-5-merous,
petals linear or obovate-spathulate; stamens free, twice as many as the petals,
filaments slender, sometimes broadened at the base, anthers small, linear;
ovary ovoid or elliptical, tapering more or less toward the base, with 3-5
locules, ovules 1 or 2 in each locule; fruits small, with the seeds imbedded in
mucilaginous pulp; seeds plump (sometimes polyembryonic in T.
trifolia).
The genus was represented from
1790 until 1925 by a single species, T. trifolia. The
discovery in the Philippines of a second species, a plant with much larger
simple leaves which Merrill named Triphasia grandifolia, broadened our
concept of the genus.
It now appears that
there is a third species, descried as Paramignya brassii from New Guinea
by C. T. White in 1926 and made the type of a new genus, Echinocitrus, by
Tanaka in 1928. This species, although obviously closely related to
the type of Triphasia, differs in a number of characters that justify
placing it in a distinct subgenus, for which Tanaka’s generic name,
Echinocitrus, is retained.
Keys to the subgenera and species of Triphasia are
presented.
Subgenus Triphasia
Subgenus
Eutriphasia Swing. in Webber & Batchelor, Citrus Indus. 1:237.
1943.
1. Triphasia trifolia
(Burm. f.) P. Wils. Torreya 9:33. 1909. Limonia trifolia Burm. f.
Fl. Ind. 103. 1768; L. trifoliata L. 1771; Triphasia aurantiola
Lour. 1790; Limonia (?) diacantha DC. 1824; Triphasia
trifoliata (L.) DC. 1824. Illus. Burmann f. loc. cit. pl. 35
(col.); Jacquin, Icon. Pl. Rar. 3:pl. 463. 1786-1793; Risso &
Poiteau, Hist. Nat. Orang. pl. 108 (col.). 1818-1822; fig. 3-14 this work.
Type.—Java (Burmann f., No. 166). Herb. Delessert, Geneva
(fide Tanaka).
Distribution.—Southeastern Asia (?), East Indian Archipelago (?); widely
cultivated in tropical and subtropical
regions.
Common name.—Trifoliate
limeberry, limeberry, or triphasia.
A
glabrous shrub or small tree with terete twigs bearing paired spines in the
axils of the leaves; leaves 3-foliolate, the terminal leaflet ovate with a
cuneate base and a rounded emarginate tip, 2-4 X 1.5-2 cm; lateral leaflets
much smaller than the terminal one (1-2-2 X 0.8-1.2 cm), broadly rounded at the
tip, cuneate at the base; petiolules very short (1.5-2 cm); petioles short
(3-5 mm), wingless; flowers appearing singly or 2 or 3 in the axils of the
leaves; peduncles short (3-4 X 1 mm); flower buds cylindrical, 10-12 X 3-4 mm;
flowers 3-merous (but with 6 stamens); sepals small (1.5-2 mm long), 3-lobed,
green, persistent; petals white, 10-13 X 3.6-4.5 mm; staminal filaments
slender, glabrous, 9-11 mm long, anthers oblong, 2 X 1 mm; disk annular or
short-cylindric; ovary ovoid or fusiform with 3 locules, each with 1 ovule,
narrowed into a slender, deciduous style with a capitate, 3-lobed stigma; fruit
ovoid or subglobose, sometimes apiculate, 1.2-1.5 cm long, dull reddish-orange
or crimson when fleshy ripe; peel with many small oil glands; seeds 1-3,
immersed in mucilaginous pulpy flesh.
Triphasia trifolia is very widely grown in all tropical and subtropical
regions as an ornamental shrub. It is also used for
hedges. The fragrant white flowers are soon succeeded on the
branches by the small dull-red berries (see fig. 3-14). The foliage is handsome, a shiny dark
green, and the plant usually makes a round-topped shrub that does not grow too
large for dooryard plantings. Triphasia trifolia has become
naturalized in certain sections of the United States, in "hammocks, fields and
cultivated grounds, coastal plain, Florida to Texas," according to Small (1933,
p. 760).
la. Triphasia
trifolia forma tetraploidea.
This
form differs from the species in having larger flower parts and thicker leaves,
and especially in having 36 chromosomes in all the somatic cells instead of 18,
the normal number for almost all plants of the orange subfamily so far
studied.
This undoubtedly tetraploid form of
the common trifoliate limeberry is like tetraploid forms of Citrus in
having somewhat thicker leaves and larger flower parts. Longley
(Traub and Robinson, 1937, p. 794) first detected the tetraploid nature of this
plant. It is one of a number of seedlings (of which all the others
are apparently diploid) grown from seeds of Triphasia trifolia collected
by Dr. J. N. Rose at Mazatlan, Mexico, in April, 1910. This is
believed to be the first instance reported of the spontaneous origin of a
tetraploid seedling of an aurantioid genus other than Citrus or
Poncirus. It is analogous to the wild-growing tetraploid form
of Fortunella hindsii, native to the mountains of southern China, of
which a diploid form is found in cultivation. Frost
(1938a) has found tetraploid mutations of several of the commonly grown
species of Citrus arising as seedlings in California.
2. Triphasia grandifolia Merr.
Phil. Jour. Sci. 26(4):458. 1925.
Type.—Mindoro Island, P.I. (Ramos, No. 40822). Herb. Bur. Sci.,
Manila.
Distribution.—Philippines:
known only from the type locality.
Common
name.—Unifoliate limeberry.
Merrill's
description of this species reads as follows: "An erect, nearly glabrous, spiny
shrub about 2 m high, the branches and branchlets slender, terete, pale
greenish when dry, the branchlets 1 to 1.5 mm in diameter,
puberulent. Leaves simple, alternate, chartaceous, oblong-ovate, 5
to 9 cm long, 2.5 to 4.5 cm wide, greenish when dry, the base broadly rounded,
narrowed upward to the somewhat acuminate apex, the lower surface paler than
the upper and conspicuously glandular-punctate; lateral nerves slender,
spreading, about 10 on each side of the midrib; petioles about 3 mm long;
stipules spiniform, rigid, stiff, 5 to 7 mm long. Flowers white,
solitary, axillary, one between each pair of spinelike stipules, their pedicels
about 6 mm long, somewhat thickened upward. Calyx 4 mm in diameter,
the tube 1 to 1.5 mm long, the three broad triangular lobes obtuse to acute,
punctate, about 2 mm long. Petals 3, imbricate, glabrous, punctate,
oblong-oblanceolate, about 1.5 cm long, 0.5 cm wide. Stamens 6,
their filaments narrowly lanceolate, tapering upward, 7 mm long, anthers
oblong, 3 mm long. Ovary 3 celled, short-stipitate, oblong, slight
thicker than the stout stipe and style, glabrous, the style 6 mm long, stigma
subcapitate. Disk truncate, about 1 mm high. Fruits
unknown."
Triphasia has been known
since 1790 as a monotypic genus, T. trifolia, the type species, was
remarkable in having three sepals, three petals, six stamens (three alternating
with the sepals and three with the petals), an ovary with three locules, and
trifoliolate leaves, so the genus name, Triphasia, was unusually
appropriate. In 1925, Merrill received from Mindoro Island in the
Philippines a second species, clearly a Triphasia, with 3-merous flowers
as in T. trifolia but with large simple leaves instead of small
trifoliolate leaves. The fruits of this interesting species are
still unknown.
Subgenus Echinocitrus (Tan.) Swing., n. comb.
3. Triphasia brassii (White)
Swing. Jour. Wash. Acad. Sci. 28:532. 1938. Paramignya brassii C.
T. White, Jour. Arnold Arbor. 7:231. 1926; Echinocitrus brassii
(White) Tan. Jour. Arnold Arbor. 9:138.
1928.
Type.—New Guinea, Papua, Rigo
(Brass, No. 817). Herb. Arnold
Arbor.
Distribution.—New Guinea: known
only from the original locality.
The original
description of Paramignya brassii (= Triphasia brassii) by C. T.
White, translated, reads as follows: "A slender bush, attaining 2 m in height,
branches drooping (fide Brass); spines numerous, single or paired,
0.8-1.5 cm long, slender, puberulous, evanescent on fast-growing large shrubs;
leaves shiny, dark green (fide Brass), with short petioles; petioles
puberulous, 2-3 mm long; varying greatly in the size of the blade, on flowering
twigs small (1.5 X 0.8 cm), usually larger (4 X 3 cm), ovate-rhomboid,
retusely acuminate at the apex, lateral veins 5-6 pairs, visible above and
below; flowers sweet-scented (fide Brass), axillary, solitary, with a
glabrous peduncle thickened toward the apex, about 1 cm long; calyx 5-lobed,
lobes broadly deltoid, about 1 cm long, margins ciliate; petals 5, obovate, 1 X
0.3 cm, with numerous oil glands outside toward the tip; stamens 10, filaments
free, 6 mm long, anthers linear, 2 mm long; ovary lageniform, short-stipitate;
stipe in dried specimens slightly ribbed; style thick, ribbed, 5 mm long,
stigma discoid; fruit red, of variable form, ellipsoid, pyriform or obovoid, 3
cm long, 1.5-2 cm diam., 5-locular, pulp scanty, seeds 1 or 2 in each locule, 1
X 0.6 cm."
The simple crenate leaves of the
type material of this species are usually broad at the base, often nearly
trangular [sic] in outline.
In
discussing this plant, which he made the type of a new genus,
Echinocitrus, Tanaka (1928c, p. 138) wrote: "…the present species
is closely allied to Triphasia in its puberulent, somewhat zigzag
branches armed with paired sharp spines, in the thin leaves without prominent
reticulation, solitary flowers with cupulate calyx and slender petals, free
filiform filaments and linear anthers, stalked ovary narrowed into a long
style, and in the few-celled bright-colored berry without
pulp-vesicles. It is, however, distinct from Triphasia
primarily in having 5-merous flowers, while both species of Triphasia
(T. trifolia and T. grandifolia) have always 3-merous
flowers. This species is far more spiny than Triphasia,
especially at the juvenile stage, and the fruit is very much larger and mostly
pointed at the base, not at the apex, the segments containing distinctly
biseriate seeds. The leaves are crenate, not entire, and the
pedicel of the fruit is very much elongated; calyx lobes are distinct and
ovoid-deltoid, not so broad and flat-triangular as in Triphasia.
The calyx lacks pubescence but is only minutely ciliate on the
margin. The appearance of the fruit is very much like an oval
kumquat (Fortunella margarita Swingle), but it lacks pulp-vesicles."
The characters mentioned above are most of them
of specific rather than generic nature. The fact that this species
is 5-merous (except that the stamens are twice as many as the petals) is not an
adequate character to separate it as a distinct genus from T. trifolia
and T. grandifolia, both of which have 3-merous flowers (except the
stamens, which are 6-merous). Such variation in the number occurs
frequently in genera belonging to the subtribe Triphasiinae, of which
Triphasia is the type genus.
XI. Pamburus
Swingle
XI. Pamburus
Swingle, Jour. Wash. Acad. Sci. 6:336. 1916. Chilocalyx Turcz.
(non Klotzsch), Bul. Soc. Nat. Mosc. 36:588. 1863. Illus. Wight,
Hook. Bot. Misc. 3:291, pl. 33. 1833; fig. 3-15 this work.
Type
species.—Limonia missionis Wight = Pamburus missionis (Wight)
Swing.
Distribution.—India: Madras
State; Ceylon; Vietnam: Annam, Cochin China; Java;
Bali.
Common name.—Pamburus.
Much-branched, very thorny shrubs or small
trees, young twigs angled, soon becoming terete; leaves simple, very thick;
petioles short, less than 1/10 the length of the leaf blade, more or less
margined but not winged, not articulated with the leaf blade; inflorescences
axillary racemes shorter than the leaves but longer than the petioles; flowers
4- or 5-merous; calyx cupulate, 4-5-lobed; petals 4 or 5, obovate or
elliptic-oblong; stamens 8-10 (twice as many as the petals), filaments free,
slender, glabrous, anthers cordate-elliptical or linear-oblong; disk cylindric,
narrower than the ovary; ovary with 5 (or sometimes 4) locules, with 2 ovules
in each locule; fruits globose (resembling a very small orange in appearance),
with a well-developed peel showing numerous oil glands, the segments usually
containing 1 or 2 seeds surrounded by a glutinous mucilage (pulp-vesicles
lacking); seeds large, thick, globose or
ovoid.
Pamburus differs from
Paramignya in its shrubby habit and lack of retrorse or hooked spines,
and in its thick leaves with rounded tips and short non-articulated petioles
without pulvinoid tips. Its relationships to the other genera of
the subtribe Triphasiinae are still obscure and should be cleared up by a
detailed study of the flowers and fruits.
Pamburus missionis (Wt.) Swingle, Jour. Wash. Acad. Sci. 6:338. 1916.
Limonia missionis Wt. Hook. Bot. Misc. 3:291. 1833; Atalantia
missionis (Wt.) Oliv. 1861; Chilocalyx ellipticus Turcz. 1863.
Illus. Wight, loc. cit. pl. 33; fig 3-15 this work.
Type.—British India, Madras Presidency, Talaar (Wight in Wallich, No.
6358-B). Herb. Kew.
Distribution.—India.
Common
name.—Pamburus.
The original description
by Wight reads as follows: "Leaves simple, elliptical-oblong, short-petioled;
racemes many-flowered, in the axils of the spines, shorter than the leaves.
"A small tree, with a round, very
branching, bushy head; the branches round, smooth, bright green, the
older ones armed with numerous large, strong, sharp thorns, the flower-bearing
ramuli flexuose, with small, straight, axillary spines.
Leaves alternate, short-petioled, oval, sometimes emarginate and slightly
crenate, oftener entire, smooth, coriaceous, dark shining green, mottled with
white spots, and perforated with numerous pellucid points.
Racemes in the axils of the spines, shorter than the leaves, somewhat
capitate. Flowers numerous, fragrant, pure white.
Calyx small, 4- or 5-lobed. Corolla 4-5-petaled,
caducous. Petals obovate. Stamens 8 or 10,
a little shorter than the petals. Filaments
subulate. Anthers erect, large in proportion to the
flowers. Pistil: germen superior, elevated on a
glandular receptacle, globular, 4- or 5-celled, with several ovules in
each cell, only one of which usually arrives at maturity.
Style cylindrical. Stigma capitate.
Pericarp a 4- or 5-celled berry; cells containing a very
glutinous mucilaginous fluid, and one roundish seed,enclosed in a thick,
firm, glandular, orange-like fruit.
"This tree
is found, rather frequently, in sandy soil near the sea-coast, flowering
towards the end of the cool season, and ripening its fruit about August or
September. The corolla falls a few hours after expansion;
hence it is difficult to procure specimens in flower. That here
figured was gathered about the beginning of March."
Swingle's description of the species given when
the genus Pamburus was established (1916a, p. 338) reads as
follows: "A much-branched shrub or small tree, armed with stout straight
spines, these 2-3 cm long, arising singly (or rarely in pairs?) on the side of
the bud in the axils of the leaves. Leaves oval, oblong-obovate or
elliptical, 6-10 cm long, 2-4 cm broad, very thick, coriaceous,
glandular-punctate, the tip rounded, sometimes slightly emarginate, the base
narrowed rather abruptly into the petiole, the margin entire, becoming gray and
apparently crenate in drying; lateral veins inconspicuous, tertiary ones not
apparent, the two faces very similar in appearance, drying to velvety
gray-green unlike those of any other member of the subfamily Citratae
[Aurantioideae]. Flowers 12-20 mm in diameter, fragrant, with small
pointed sepals and 5 or 4 white obovate caducous petals about 1 cm
long. Pistil about 1 cm long. Fruit about 2.5 cm in
diameter, orange-colored when ripe, with a thick peel dotted with oil glands,
5-4 celled, the cells containing 1 or 2 seeds surrounded by a sticky gum."
This remarkable plant has leaves unlike those of
any other member of the orange subfamily (see fig. 3-15). In drying they become pale gray-green and
show what seems to be a fine-textured, velvety surface. As it grows
commonly in the dry region of Ceylon and in the flat, sandy coastal lands of
Madras State in India, it may prove to be the fourth genus of the orange
subfamily to show somewhat xerophytic structure, the other three being
Oxanthera of New Caledonia, and Microcitrus and
Eremocitrus, both of the semiarid regions of northeastern Australia.
The wood of P. missionis is used in India
for making furniture. The species should be tested as a rootstock
for Citrus with Oxanthera fragrans and Eremocitrus glauca
in the hope of finding rootstocks able to grow in soils with heavy content of
mineral salts. Pamburus missionis was introduced into the
United States several decades ago.
In 1941, a
five-year-old tree of this species growing at the Plant Introduction Garden in
Coconut Grove, Florida was about 2.5 meters high and 1.5 to 2 meters wide, with
a very dense top composed of exceedingly numerous, short, divergent, very leafy
twigs with stout, very sharp spines, 1.5 to 4 cm long, attached nearly at right
angles to the twig every 2 to 4 cm. The younger branches, the
spines, and the more or less exposed under surfaces of the leaves were all of
the same vivid green color, close to grass green (Ridgeway, 1912, pl.
17). The upper surfaces of the leaves, also visible in large part,
were a slightly darker green, near Hay’s green (Ridgeway, 1912, pl.
6). The foliage was not only vivid green, but perfectly smooth and
glossy, and showed no traces of injury from insects or diseases.
This plant seems admirably suited for making beautiful hedges of moderate
height, since it stands pruning well.
Anatomical study of freshly gathered leaves shows that the upper surfaces have
two layers of palisade cells and the under surface one layer. This
doubtless explains the almost identical appearance of the upper and lower
surfaces of the leaves and the almost complete masking of the veinlets.
The plants grown at Coconut Grove thrived on
rather poor, sandy soil that thinly covered a subsoil composed of porous
limestone rock. The pamburus seems to withstand nearly as much cold
as the commonly cultivated species of Citrus.
GROUP C. THE LUVUNGA
GROUP
The two closely related genera Luvunga and
Paramignya, with a total of twenty-seven species, are both clambering
woody lianas that hold fast to the branches of trees by means of retrorse or
sometimes strongly recurved spines. The leaves of Luvunga
are large, apparently palmately trifoliolate, with very long wingless petioles;
those of Paramignya are medium-sized, unifoliolate, with short
petioles. Both genera have the petioles articulated at both ends
(at the junction with the leaf blade above and with the twig
below). These joints at both ends of the petioles are more or less
swollen and pulvinoid, which permits each leaf to take up the best possible
position to catch the light. The ovaries are 2- to 5-locular and
each locule has two ovules. The fruits, so far as known, do not
contain pulp-vesicles.
The fruits both of
Luvunga and of the typical species of Paramignya show a striking
dimorphism of the oil glands, those in the peel being small, those in the
deeper tissues between the peel and the locule cavities being much
larger. Possibly the deeper glands may produce not oil but mucilage
that ultimately flows into the locule cavities about the seeds as the fruits
ripen. Only the study of fresh living fruits can decide the
matter.
It is highly probable that
Luvunga and Paramignya are descended from a common ancestral plant
with pinnate or trifoliolate leaves. When Luvunga plants bear
unifoliolate leaves, they have very much shorter petioles than do the normal
trifoliolate leaves and, in consequence, look much like normal
Paramignya leaves. The flowers of both Luvunga and
Paramignya look much like those of Citrus except for having fewer
stamens (only twice as many as the petals instead of the four or more times as
many found in Citrus). The fruits look much like miniature
lemons or limes, and, like ordinary Citrus fruits, have a peel thickly
dotted with oil glands.
Abundant material of
the genera Luvunga and Paramignya studied by Tillson (1938, pp.
11, 12, 24) shows that in the development of the flower the petal midrib, as it
diverges from the axis, carries, fused with it, the lateral sepal traces, an
unusual type of floral vascular anatomy, characteristic of the Primitive Citrus
Fruit Trees of the subtribe Citrinae, of the species of the subgenus
Citrus (comprising all the commonly cultivated citrus fruit trees), and
of Swinglea (see Tillson and Bamford, 1938, p. 791).
It is possible that more thorough study of the
developing fruits will show that Luvunga and Paramignya are so
closely related to the Primitive Citrus Fruit Trees that they should be
transferred out of the subtribe Triphasiinae to the subtribe
Citrinae. In the meantime, as the highest group (C) of the Minor
Citroid Fruit Trees, the Luvunga group is placed next to the Primitive
Citrus Fruit Trees (group A) of the subtribe Citrinae.
XII. Luvunga Buch.-Ham.
XII. Luvunga6 (Lavanga) Buch.-Ham.
Wall. Cat. 217. 1831; Wt & Arn. Prodr. 1:90. 1834. Lampetia
Roem. Syn. Hesper. 1:42. 1846.
Type
species.—Luvunga scandens (Roxb.) Buch.-Ham.
Distribution.—India, Ceylon, Burma,
Thailand, Indo-China, Malay Peninsula, Sumatra, Java, Borneo, Philippines, and
New Guinea.
Common name.—Trifoliate
liana-limes (or luvunga).
Woody lianas with
single, recurved axillary spines; leaves 3-foliolate with long, wingless
petioles; leaflets oval to lanceolate, acute or acuminate at the apex, entire,
often slightly revolute on the margins; inflorescences dense axillary racemes
or panicles; flowers 4- or 5-merous; calyx cup-shaped, 4-5 lobed; petals 3-5,
linear or lanceolate, thick; stamens 6-10 (rarely fewer), with free, or more or
less connate, filaments and linear anthers; ovary elongate-ovoid, seated on a
columnar gynophore, locules 2-4, with 2 collateral or superimposed ovules in
each locule; style gradually merging with the ovary, thick, sooner or later
caducous, stigma capitate; fruits ellipsoid or globose, rind thick; seeds 1-3,
imbedded in mucilaginous matter (pulp-vesicles not
present!).
Luvunga includes about
twelve species which are very similar in general appearance. All
the species are woody vines that cling to forest trees by means of strong,
recurved spines in the axils of the leaves. They also agree in
having trifoliolate leaves (occasionally unifoliolate or difoliolate), borne on
very long, wingless petioles, and in bearing crowded clusters of strongly but
agreeably scented, white, orange-like flowers in the leaf axils.
The fruits are small, globose or ovoid, usually rough-skinned, and yellow in
color; they somewhat resemble miniature lemons or oranges in appearance but
contain no juice. Instead they are filled with a sticky
mucilaginous substance in which the few medium-sized seeds are imbedded.
The species of Luvunga are difficult to
distinguish from a study of herbarium specimens and need critical study
badly. Only ten species are well enough known to be keyed out here, but two more of doubtful validity are
added. Few genera in the orange subfamily show so little diversity
among the species as does Luvunga, but this genus is not closely related
to any other of the Minor Citroid Fruit Trees except Paramignya, which is
also a genus of woody vines that hold fast to the branches of trees with their
reflexed spines. Luvunga differs from Paramignya
chiefly in having an unusual type of trifoliolate leaves on long, stiff
petioles. Occasionally unifoliolate leaves with short petioles are
produced by species of Luvunga; such leaves bear a close resemblance to
the characteristic leaves of Paramignya. It is possible, even
probable, that the genus Paramignya arose from an aberrant species of
Luvunga which produced only unifoliolate leaves.
The fruits of Luvunga show almost exactly
the same structure as those of Paramignya. They have small oil glands in
the peel and much larger ones in the tissue between the peel and the locule
cavities. This dimorphism of the oil glands (possibly oil glands
and mucilage glands?) is very evident in fruits of L. sarmentosa growing
in the Botanic Garden at Buitenzorg, Java, collected by C. S. Sargent, October
16, 1903, now in the herbarium of the Arnold Arboretum (Tanaka, Det. No.
A-325), and in L. crassifolia collected in Sumatra by H. S. Yates (No.
254) in 1922, now in the herbarium of the University of California, sheet
226,095 (Tanaka, Det. No. C-254).
All the
species of Luvunga are woody vines that clamber high on trees of
tropical evergreen forests. The fruits are small, 1 to 3 cm in
diameter, globular or lemon-shaped, with a rough peel and a few medium-sized
seeds. These luxuriant vines, bearing thousands of densely
crowded, fragrant white flowers, may prove useful as ornamentals in southern
Florida, Puerto Rico, and other rainy tropical regions.
1. Luvunga scandens (Roxb.)
Buch.-Ham. Wall. Cat. 6382. 1831. Limoniascandens Roxb. Fl. Ind.
2:380. 1832. Illus. Hooker, Bot. Mag. 86:pl. 4522 (col.).
1850; Paxton, Fl. Gard. 1:109. 1850-1851; Lemaire, Jard. Fleuriste
1:pl. 75 (col.). (copied exactly from Hooker, loc. cit.) 1851;
Schnizlein, Iconogr. 3:pl. 224. 1857-1865; Pierre, Flore Forest.
Cochinch. 4:pl. 288A. 1893; Guillaumin, in Lecomte, Fl. Gén. Indo-Chine
1:667, fig. 71(1-2). 1911; Basu, Ind. Med. Pl. pl. 194. 1918; fig. 3-16 this work.
Type.—British India, hills about Silhet (Roxburgh). Herb. Hort.
Bot., Calcutta.
Distribution.—India,
Burma, Laos, North Vietnam, South Vietnam, Thailand; also occurs in Sumatra and
Borneo (fide Tanaka).
Common
name.—Indian liana-lime (or luvunga).
Twigs with long, acute, slightly recurved thorns; leaves variable; petioles 5-13
cm; leaflets slender, elliptic-oblong or oblanceolate, acute or acuminate at
the tip, cuneate at the base, venation inconspicuous; inflorescences small,
axillary cymes; flowers about 1.5-2 cm diam.; calyx with 4 short, truncate
lobes; petals 4, white, fleshy, recurved, 12.5-18 mm long and 2-6 mm wide;
stamens 8, glabrous, with the filaments connate nearly to the tips; ovary with
3 or 4 locules, ovules 2 in each locule; fruits oblong, about 25 X 20 cm ("size
of a pigeon's egg"), rather smooth; seeds oval, somewhat pointed, integument
thin, monoembryonic cotyledons oblong, green.
This, the type species of the genus, is easily recognized by the smooth,
entirely hairless staminal filaments that are fused together nearly to their
tips into a tube and by the long, slender, pointed leaflets. It is
native from northeastern India through northern Burma, Laos, Cambodia, and
North and South Vietnam. Luvunga scandens is the best-known
species of the genus, as it has been very well illustrated by Hooker and
Pierre. It can be easily grown in a greenhouse but, according to
experiences in Europe, it does not flower unless given opportunity to climb
high, as, for instance, in a palm house.
2. Luvunga crassifolia Tan. Med. Rijks Herb. Leiden
69:8. 1931.
Type.—Malacca
(Maingay, No. 285). Herb. Kew.
Distribution.—Malay Peninsula, Borneo, Sumatra.
Tanaka's original description, translated, reads
as follows: "Branches thick; spines curved; petioles short, somewhat thick;
leaves very thick, obovate or oblong-obovate, rounded at the apex, frequently
emarginate, often apiculate, obtuse at the base, margins slightly revolute,
shiny above, with distinct veins, dull below, minutely verrucose.
Inflorescence many-flowered; flowers large; calyx deeply cup-shaped; filaments
connate, puberulous. Fruit large, tubercular, foveolate."
This may be supplemented by the following notes
Swingle was able to make from a specimen in the herbarium of the Arnold
Arboretum, identified by Tanaka as L. crassifolia (Tan. ident., No.
A-328; coll. by H. S. Yates, No. 766, east coast of Sumatra).
This specimen has rather thick, semicoriaceous
leaves, with 12-15 pairs of lateral veins visible on both surfaces, more
conspicuous below, arising at an angle of 45°-55° with the midrib; terminal
leaflets 21-22.5 X 7.5-8 cm, lateral leaflets 16.5-18.5 X 6-6.5 cm; petiole
11-14.5 cm long. An inflorescence shows 6 fruits with a persistent,
cup-shaped calyx, 7-9 mm wide, 2.5-3 mm high, with 5 (?) blunt, short lobes;
pedicels 6-8 mm long, articulated with the short lateral branches (3-5 mm long)
of the inflorescence; fruits (dried herb. spec.) fusiform, apex acute, with
apparently persistent style, base abruptly rounded.
This species differs from L. scandens in
having blunter, thicker leaves, with evident venation, and in having pubescent
filaments. L. crassifolia was evidently included in L.
scandens in many of the older floras of the Malayan region.
3. Luvunga nitida Pierre,
Flore Forest. Cochinch. 4: text to pl. 288B. 1893. Illus.
Pierre, loc. cit. pl. 288B
(1-8).
Type.—French Indo-China,
Cambodia, Province of Tran (Pierre, No. 3876). Herb. Mus. Hist.
Nat., Paris.
Distribution.—South
Vietnam: Pell Mountains in Chaudoc Province; Cambodia: Tran Province (the
original localities cited by Pierre; not known elsewhere).
Leaflets elliptic or oblong, abruptly acuminate,
obtuse or acute at the tip, usually obtuse at the base (11-20 X 5-8 cm), having
about 10 pairs of widely spaced, faint nerves; petioles 7-8 cm long; flowers
4-merous, peduncles 3-4 mm long; sepals 4, fused together, ciliate on the
margins; petals 4, oblong, rounded and blunt-pointed at the tip, reflexed when
fully open, stamens usually 6 or 7, filaments glabrous, slender, free except at
the very base, where they are connate, anthers oblong; ovary, with 3 or 4
locules, seated on a disk 1 mm high, with 2 superimposed ovules in each locule;
style rather thick, 4.5-5 mm long, summit expanded into the truncate stigma;
fruit unknown.
This species was considered a
synonym of L. scandens by Tanaka (1930a, p. 52).
However, the free stamens and the broad leaflets, bluntly rounded at the base
with only about ten pairs of faint veins, as well as the style gradually
expanded into the truncate stigma, show that it is quite different.
It is much more closely allied to L. eleutherandra, from which it
differs in having glabrous, filiform staminal filaments and larger leaflets with
obtuse bases.
4. Luvunga
eleutherandra Dalz. Hook. Kew Jour. Bot. 2:258. 1850. Illus.
Engler, Die Nat. Pflanzenfam. 3(4):189, fig. 109,M-R (fls.).
1896; ibid. 19a:323, fig. 147,M-R. 1931; Talbot, Forest
Fl. Bomb. Presid. 1:197, fig. 120 (lvs. and fts. [sic]). 1900.
Type.—British India, Bombay
Presidency, Syhadree Mountains (Dalzell, No. 54). Herb.
Kew.
Distribution.—India, Ceylon (also
reported, probably erroneously, from Malay Peninsula, Sumatra, and Java).
Branches flexuose, with sharp, somewhat
deflexed, nearly straight spines, 1.5-2 cm long; leaflets 7.5-13 X 3.5-6.5 cm,
elliptic-oblong or obovate, shortly acuminate, base usually acute; petiole
2.5-7.5 cm long; flowers small, 4-merous; calyx cup-shaped, almost entire or
obscurely 4- or 5-lobed, truncate, puberulous; petals 4, elliptic-oblong,
reflexed, 6-7 mm long; stamens 8, shorter than the petals, filaments free,
dilated, finely pubescent; style short, stout; fruits ellipsoid, small, 2.5-3.5
X 1.5-2 cm, peel rough and pitted.
This
small-leaved and small-flowered species is found, according to C. E. C. Fischer
(1921), in the western part of peninsular India from Konkan southward to the
Anaimalai Hills in the Coimbatore district in evergreen forests above 3,000 ft.
altitude (between Lat. 10° 13' and 10° 32' N.).
5. Luvunga angustifolia
(Oliv.) Tan. Bul. Soc. Bot. France 75:711. 1928. Luvunga eleutherandra
Dalz. var. angustifolia Oliv. Jour. Linn. Soc. Bot. 5(2):44.
1861.
Type.—Ceylon (Walker and
Thwaites, No. 1195).
Distribution.—Ceylon.
Branches with spines
like those of L. eleutherandra but slightly shorter (about 2.5 cm);
leaves 3-foliolate with petioles 3.4-6.5 cm long; leaflets very variable,
oval-lanceolate or linear-lanceolate, shortly acuminate at the apex, entire,
7.5-20 cm long; filaments free, subulate, glabrous (instead of finely pubescent
as in L. eleutherandra); fruits 2 cm diam.; globular (instead of
ellipsoid as in L. eleutherandra).
This
Ceylon species as described by Trimen, (1893, p. 224) and Tanaka (1928b,
pp. 711-12) appears to be fairly distinct from L. eleutherandra.
A careful comparison should be made of the plant of western peninsular India
on which Dalzell based his L. eleutherandra in 1850 and of the Ceylon
plant on which Oliver established his variety beta (angustifolia)
in 1861. Both the species of western peninsular India and that of
Ceylon were said by Talbot (1909, p. 197) to grow at altitudes of from 3,000 to
5,000 feet in evergreen forests. L. eleutherandra occurs only
about 200 miles northeast of the mountain region of south-central Ceylon where
L. angustifolia is found. L. angustifolia is said to
show a tendency to dioecism. Thwaites stated (1864, p. 48): "…this
species would appear to be dioecious, as in some plants the ovaries of all the
flowers are entirely without ovules."
6. Luvunga sarmentosa (Blume) Kurz, Jour. Asiat. Soc. Bengal
39(2):69. 1870. Triphasia sarmentosa Blume, Bijdr. Fl. Nederl.
Indie 152. 1825.
Type.—Java, Salak
(Blume). Rijks Herb., Leiden.
Distribution.—Java; Sumatra; Borneo; Malaya; southern Burma: Tavoy in
Tenasserim.
Leaves 3-foliolate, leaflets
usually small, about 12.5 cm long (but up to 30 cm long on young plants,
according to Koorders, 1912, p. 426), obscure venation, elliptical or obovate
(young leaflets sometimes linear, acuminate, as narrow as 30 X 3 cm), abruptly
acuminate at the tip, sometimes short-caudate, but the very tip bluntly
rounded, acute or broadly rounded at the base; flowers small, with free, very
pubescent filaments; calyx 2-3 mm diam., with 5 triangular, minutely pubescent
acutish lobes; fruits small.
In spite of
Kurz's recognition of this plant in 1870 as a species of Luvunga distinct
from L. eleutherandra, it was not considered by other taxonomists to be a
distinct species until Tanaka (1931b, p. 9) published very helpful notes
on the species of Luvunga found in the Dutch East Indies (now West Irian,
Indonesia). He examined in the Leiden herbarium the type specimens
of Blume on which L. sarmentosa was founded and was able to distinguish
it from L. eleutherandra of western peninsular India and from L.
angustifolia of Ceylon.
7.
Luvunga borneensis Hochr. Cat. Hort. Bogor. 1:41. 1904; and
(simultaneously?) in Pl. Bogor. Exsic. 19.
1904.
Type.—Borneo, Sambus
(Hochreutiner). Herb. Hort. Bot., Buitenzorg, Java (now Kebun Raya,
Bogor, Java).
Distribution.—Known only
from the original locality, but it is cultivated in the Botanic Garden in Bogor,
Java.
[After Hochreutiner]: "Differs from
L. eleutherandra [probably L. sarmentosa is intended] having
corolla and especially the calyx (5 mm high and 5 mm wide) larger; flowers
5-merous [instead of 3-4-merous]; stigma capitate and much larger; flower buds
acorn-shaped; inflorescences fewer-flowered. It differs from L.
Motleyi in having petioles 7.5-15.5 cm long; petioles 8-10 mm long;
leaflets 9-16.5 cm long and 5-7.5 cm wide; flowers in cymes no denser than in
L. eleutherandra [L. sarmentosa?]."
Unfortunately, no additional information has
been published since 1904 and no additional collections of this plant have been
made in Borneo. Tanaka did not see the type specimen in the
herbarium of the Botanic Garden at Buitenzorg (now Bogor), Java, nor did he
report on Hochreutiner's authentic specimens of this species distributed in his
Plantae bogorienses exsiccatae, No. 30, in 1904. It has been
possible to examine (in Washington, D.C.) an authentic flowering specimen of
this species distributed in the exsiccatae by Hochreutiner as well as a
specimen from another plant growing alongside in the same botanical
garden. They show the following characters: Spines on sterile
shoots about 2 to 2.5 cm long, very strongly recurved and very sharp, but none
on the flowering shoot; leaves trifoliolate; petioles 2.5 to 8.5 cm (usually 5
to 8 cm) long; leaflets broadly elliptical, abruptly acuminate at the apex,
acumen blunt on the authentic flowering specimen and acute on the sterile
plant, broadly cuneate or rounded (on small leaves) at base, with 10 to 14
pairs of parallel veins scarcely visible on the upper surface but very evident
on the under surface in dried specimens; inflorescences axillary, composed of 1
to 4 short (3 to 4 cm), divaricately branched cymes; pedicels 3 to 4 mm long;
calyx cup-shaped, very finely pubescent, 3.5 to 5 mm wide and 3 to 4 mm deep,
with five broadly rounded lobes, finely ciliate on the margins; corolla and
stamens lacking; pistils 7 to 9 mm long; ovary slender, finely pubescent.
8. Luvunga motleyi Oliv. Jour.
Linn. Soc. Bot. 5(2):44. 1861.
Type.—Borneo, Banjarmassing (Motley).
Distribution.—Borneo, Sumatra.
[After
Oliver, 1861, p. 44]: Branches slender, glabrous; petioles of the size of a
crow's quill, 10-12 in. [25.5-30.5 cm] long; petiolules sometimes 8-10 lin.
[16-20 mm] long; leaflets obovate-lanceolate, entire or slightly
undulate-crenate, 9-18 in. [23-46 cm] long, 4-6 in. [10-15 cm] wide; flowers on
short pedicels, in dense, depressed, axillary cymes; calyx somewhat cupulate,
with 5 small, triangular lobes or scarcely lobed; petals 5 (?); stamens 10,
free; ovary with 3 or 4 locules.
Tanaka
(1931b, p. 9) noted that this species has extremely large leaflets with
distinct venation, large flowers with shallow calyx, and free, very pubescent
filaments.
9. Luvunga
philippinensis Merr. Phil. Jour. Sci. Bot. 3:233.
1908.
Type.—Philippines, Mindanao,
Zamboanga (Whitford and Hutchinson, No. 9104). Herb. Bur. Sci.,
Manila.
Distribution.—Philippines:
Mindanao Island; Borneo.
[After Merrill]: A
climbing vine; petioles 9-13 cm long; petiolules stout (5 mm); leaflets 15-25 X
6-10 cm, subcoriaceous, oblong to oblong-elliptical, apex acuminate, the
acumen blunt or retruse, base acute, margins entire, with 8-10 pairs of
irregular nerves, somewhat prominent beneath, anastomosing, "inflorescences of
small, usually 3-flowered, racemose cymes, borne on the branches below the
leaves or in the leaf-axils"; pedicels 4-5 mm long; flowers white, fragrant;
calyx cup-shaped, about 4 mm wide, truncate or very obscurely 5-toothed, its
stipe 2 mm long; petals 3 or 4, oblong, obtuse at apex, 9-10 X 3.5-4.5 mm;
stamens free, filaments short, swollen, glabrous, about 6 mm long; ovary with 4
locules, oblong, quadrangular in cross section, about 3 mm long, 1.2 mm thick,
scarcely narrowed into the stout style, nearly equaling the ovary in length;
stigma capitate.
Tanaka (1931b, p. 9)
found that Luvunga philippinensis differs from his L. crassifolia
in having indistinct venation, small flowers, free pubescent filaments, and
large fruits borne on a small calyx.
The
original description was based on specimens with spineless twigs, such as are
not uncommon in fruiting branches. Undoubtedly rapidly growing
young branches of this species will be found to bear the retrorse (but not
coiled) spines characteristic of this
genus.
Luvunga philippinensis is well
distinguished from L. papuana (the only species known to the south and
east some 3,600 km distant in northeastern New Guinea), which has smaller
acuminate leaflets, longer style, ovary with 2 to 4 locules, and much more
strongly recurved spines, sometimes making a complete circle or even more!
It also differs from the species which occur
from 800 to 1,600 km to the southwest, in Borneo; (1) from L. borneensis,
in having 3- or 4-merous flowers, with smaller calyx, longer petioles, and
larger leaflets; (2) from L. motleyi, in having shorter petioles, smaller
leaflets, and smaller flowers with glabrous filaments; and (3) from L.
crassifolia, in having inconspicuous, irregular lateral veins, smaller
flowers, glabrous filaments, and leaflets acute at the base.
10. Luvunga papuana Lauterb.
Bot Jahrb. 55:260. 1918. Illus. Lauterbach, loc. cit.
55:261, fig. 7,A,K; Engler, Die Nat. Pflanzenfam.
19a:324. 1931.
Type.—Northeastern New Guinea, Mai River, Sepik (Ledermann, No.
7392). Cotype: Pfingstberg, Sepik (Ledermann, No.
7505).
Distribution.—New Guinea.
[After Lauterbach's descriptions and figures]:
Branches terete, with axillary, very strongly reflexed, slender, sharp spines
coiled in a spiral manner (sometimes more than one turn!); leaves 3-foliolate or
rarely 1-foliolate; petioles short (about 10 mm on 1-foliolate leaves; 8-17 cm
on 3-foliolate leaves); leaflets 10-18 X 6-9 cm, broadly lanceolate,
obtusely-acuminate at the tip, acute and decurrent at the base, subcoriaceous,
shiny above, dull below, margins entire, revolute, with about 9 pairs of
inconspicuous lateral nerves; inflorescences fascicled, many-flowered, axillary
cymes, 2-4 cm diam.; pedicels 5 mm long, calyx (in the flower bud) 1.3 mm
diam.; petals 3-4, 6 X 3 mm; stamens 8, 5 mm long, with glabrous filaments
1-1.5 mm long in the flower bud; ovaries ovoid, lageniform, glandular,
2-locular, with 8-10 vertical furrows (probably between rows of oil glands);
style thick, 4 mm long, merging gradually into the ovary; stigma somewhat
capitate ; fruits globose, about 12 mm diam., rind thick, granular, white;
seeds 1, 8 mm long, with a membranous testa.
This species, which is the southeastern-most species of the genus and the only
one found so far in New Guinea, is strikingly different from the others in
having slender, sharp, very strongly recurved spines (shown by Lauterbach as
arching downward to touch the stems below). The ovary is shown as
having glands apparently arranged in eight or ten vertical lines marked by
slight ridges on the ovary. The fruits are the smallest known in
the genus, being only about 12 mm in diameter. Both the description
and the illustration show that some of the leaves are unifoliolate and very
short-petioled (as short as 1 cm). This species seems clearly
distinct from all others known in the genus.
A
plant apparently belonging to this species was collected for the 1936 Richard
Archbold Expedition to New Guinea by L. J. Brass (No. 7425, August, 1936) at
Oroville camp, Fly River, in Papua, southeastern New Guinea. A
specimen in the herbarium of the Arnold Arboretum has Brass’s notes on the
label which read: "fruits white, obovate, about 3-5 cm more or less long and 3
cm diam." The strongly recurved spines and coriaceous leaves,
shiny-green above, make it very probable that this plant is L. papuana
in spite of its large fruits, and make it likely also that the small fruits
mentioned by Lauterbach in the original description were
immature.
IMPERFECTLY KNOWN SPECIES
11. Luvunga calophylla Kurz, Jour. Asiat. Soc. Bengal
39(2):69. 1870.
Type.—Bangka
Island (near Sumatra) at Jébús (Teijsmann, No. 3223). Herb. Hort.
Bot., Buitenzorg, Java (now Kebun Raya, Bogor,
Java).
Distribution.—Known only from
the type locality.
[After Kurz]: Leaves
3-foliolate; petioles 20-23 cm long; leaflets 25-30 X 4 cm, obovate-lanceolate,
shiny on both surfaces, lateral nerves conspicuous; flowers in short, glabrous
cymes; calyx large, 5-lobed, truncate, glabrous; petals, styles, etc., lacking;
fruits (immature) oblong or ovate-oblong, showing the base of the style,
"vesiculose-papillose." Kurz said of it: "A very distinct species
with leaves much resembling those of Zanthoxylum euneurum Miq. [=
Evodia euneura Miq.]."
The absence of
flowers has prevented the accurate placing of this species. Kurz
knew Luvunga well and his opinion is of value. L.
calophylla is carried in the Index Kewensis (Hooker and Jackson,
1895-1938) as a valid species.
12.
Luvunga tavoyana Lindl. ex Oliv. Jour. Linn. Soc. Bot. 5(2):44.
1861.
Type.—Southern Burma, Tenasserim,
Tavoy ("W. G[riffith]," Wall. Cat., p. 217, No.
6383).
Distribution.—Known only from
the original collection.
[After Oliver, 1861,
p. 44]: Leaves with petioles 7.5-15 cm long; leaflets 10-20 cm long,
oblong-elliptical; flowers in short fascicles, or panicles, 1.2-5 cm long;
stamens free or slightly united at the base; ovary with 3 or 2
locules. "It occurs both unarmed and with strong recurved axillary
spines."
Oliver did not consider this to be a
good species but rather a connecting link between L. scandens and L.
eleutherandra that might warrant their union under one species.
Such a union seems very improbable in the light of our present knowledge of
Luvunga. L. tavoyana should be compared critically
with L. calophylla and L. nitida. It cannot be placed
with any certainty at present.
XIII. Paramignya
Wight
XIII. Paramignya
Wight, Ill. Ind. Bot. 1:108, 110, pl. 42. 1840. Atlantia Guill.
(pro parte, non Corrêa), in Lecomte, Not. Syst. 1:176.
1910.
Type species.—Paramignya
monophylla Wt.; native to British India, Ceylon, and southern
Burma.
Distribution.—India; Ceylon;
Burma; southern China: Hainan Island; Laos; Cambodia; North Vietnam; South
Vietnam; Thailand; Malay Peninsula; Sumatra; Sumbawa; Borneo; Philippines;
northeastern Australia.
Common
name.—Unifoliate liana-limes (or paramignya).
Woody vines clambering up trees and shrubs by
means of recurved spines; leaves 1-foliolate; petiole usually showing a
pulvinoid, more or less swollen segment articulated with the leaf blade above,
and the rigid portion of the petiole is also articulated with the twig below;
flowers white, fragrant, often large, 4-5-merous; stamens twice as many as the
petals, usually with elongate-linear anthers; disk an elongated columnar
gynophore; ovary with 3-5 locules, 1 or 2 ovules in each locule; fruits globose
or obovoid, often being narrowed toward the base, with a thick peel, seeds
rather flat.
As here used, Paramignya
is a genus containing about twelve very closely related species, often
difficult to distinguish from a study of herbarium material. Two
species native to Thailand, P. surasiana and P. rectispinosa, and
one native to the island of Hainan in extreme southeastern China, P.
confertifolia, have very small ovaries, much shorter than the styles, but
otherwise they are very similar to the typical species of the genus.
Besides the twelve typical species of
Paramignya there are three aberrant species that may possibly prove upon
study of good material not to belong to the genus Paramignya.
One of these species, P. lobata, a native of the Malay Peninsula, has
prominently 2- to 5-lobed fruits, usually broader than long and bright red in
color when ripe. This species has very short retrorsely curved
spines and is evidently a woody climber. P. cuspidata, also
native to the Malay Peninsula, similarly has very short spines but apparently
they are not retrorse; it is not a high-climbing vine but rather a spiny
straggling shrub that probably clambers over low trees, shrubs, and other
vegetation. It sometimes has recurved spiny lateral twigs on the
branches that might aid in clambering over other plants. The third
aberrant species is P. trimera, which has a wide range from northeastern
Australia and Timor Island to the islands of the southern Philippines and
Indonesia.
The fruits of the typical species of Paramignya have small
or medium-sized (0.1 to 0.6 mm) oil glands in the peel, and larger, sometimes
much larger, ones (0.8 to 2 mm) below the peel in the mesocarp or even
sometimes in the endocarp, not far from the locule cavity. In P.
longipedunculata there are very commonly two layers of oil glands, one
composed of very small (0.1 to 0.5 mm) glands in the peel and another
(sometimes two layers deep) composed of much larger (0.5 to 1.5 or 2 mm)
globular or ellipsoid glands in the mesocarp with the longer diameter
tangential. In this species, the two kinds of oil glands (the
larger ones perhaps mucilage glands) are separated by a layer containing
numerous small vascular bundles.
The typical
species of Paramignya are undoubtedly closely related to
Luvunga. Both of these genera include woody vines clambering
high into trees and holding fast by means of recurved spines, and both genera
have fruits with dimorphic oil glands. Luvunga has
trifoliolate leaves with very long petioles, whereas Paramignya has
unifoliolate short-petioled leaves. There are other minor
differences such as the straight twigs (with few internodes) of Luvunga
and the more or less zigzag twigs of Paramignya; also the presence of a
well-defined pulvinus, at the base of the leaf in Paramignya, which
twists so as to place the leaf in the proper position to catch incident
light.
Sometimes Luvunga produces
unifoliolate leaves and then the petioles are very much shorter, not very
different in length from those of some of the larger-leaved species of
Paramignya, for example, P. andamanica. Probably these
two genera were derived from some common ancestor. The aberrant
species P. lobata and P. cuspidata may, however, be related to
Pamburus.
The species of
Paramignya are difficult to distinguish unless cognizance is taken of
all available characters. Unfortunately, many of the published
descriptions merely distinguish between the few species growing in a single
region; this does not permit accurate comparison of species growing in distant
regions. For the present it is easiest to key out separately the species of the four principal regions
where this genus is native.
1.
Paramignya monophylla Wight, Ill. Ind. Bot. 1:108, pl. 42. 1840.
Atalantia correae Guill. 1910. Illus. Wight, loc. cit. pl.
42; Engler, Die Nat. Pflanzenfam. 3(4):191, fig. 111,A-B.
1896; ibid. 19a:327, fig. 150,A-B. 1931; Talbot,
Forest Fl. Bomb. Presid. 1:200, fig. 122. 1909; Guillaumin, in Lecomte,
Not. Syst. 1:176, fig. 8(1). 1910; Swingle, in Bailey, Stand. Cycl. Hort.
5:2473, fig. 2763. 1916.
Type.—British India, Courtallum (Wight, No. 115). Herb. Univ.
Glasgow (fide Tanaka, 1930b, p.
235).
Distribution.—India: western
peninsula from Konkan northeast to Sikkim, Bhutan, Khasiak; Burma: Tenasserim;
Ceylon.
Common name.—Indian
paramignya.
Leaves oblong or elliptic, rarely
obovate, 6-12.5 X 2-4 cm, apices abruptly acute, sometimes acuminate, base
rounded; petioles 12-20 mm long; flowers solitary or in few-flowered axillary
clusters, large (25 mm diam.), 5-merous, fragrant; calyx cupular, with 4-5
broad, obtuse lobes; petals 4-5, lanceolate-oblong; stamens 8 or 10, free,
filaments slender, compressed below, subulate above, glabrous or pubescent;
pedicels slender, 6-10 mm long; ovary slightly furrowed, pubescent or glabrous,
with 3-5 locules, usually with 2 ovules (sometimes 1) in each locule; style
pubescent below, glabrous above; fruit ovoid or obovoid, pubescent or almost
glabrous, slightly angled and sometimes compressed, 2.5-3.5 X 2.5-3.3 cm,
yellow, odorous, sometimes only 1-seeded; seeds compressed.
This species, the type of the genus, has never
been adequately studied or illustrated. The petioles are not
clearly articulated between the leaf blade and the twig, but the upper portion
is pulvinoid. The fruit was said by Wight in the original
description to be "pomaceous! endocarp 5-angled, between coriaceous and fleshy,
surrounded by cellular pulp and rind, one- (always?) celled by the rupture and
absorption of the partitions." He also stated that the fruit is
"obtusely 5-angled and furrowed between, clothed with short matted
pubescence." Oliver (1861, p. 42) described the ovary as "often
slightly furrowed."
2.
Paramignya scandens (Griff.) Craib, Fl. Siam. Enum. 1:235. 1926.
Citrus scandens Griff. Not. Ad. Pl. As. 4:495. 1854; Paramignya
citrifolia Oliv. Jour. Linn. Soc. Bot. 5(2):42 (excl. synonymy! [non
Limonia citrifolia Roxb.]). 1861; P. griffithii Hook. f. Fl. Brit.
Ind. 1:510. 1875; Atalantia griffithii Guill. in Lecomte, Not.
Syst. 1:183. 1910. Illus. Griffith, loc. cit. pl. 587, fig.
1.
Type.—Northern Burma, "Serpentine
Mines," Mogung (?) Valley (Griffith, No.
519).
Distribution.—Northeastern India:
Assam State; northern Burma; Thailand.
The
original description by Griffith reads, translated, about as follows: "A spiny
climbing shrub; spines smaller on twiglets or on inflorescences; twigs,
petioles, and leaves pubescent; flowers white like the stamens, fragrant (with
a suave odor), petals tinted green outside above the middle.
Habitat: In wooded hills near the Serpentine Mines, toward Burma.
"A very distinct species, rare in upper Assam;
common in the valley of the Mogoung, especially four leagues toward Serpentine
Mines."
Griffith (l.c., vol. 4, pl. 587) shows
a flowering twig, 24 cm long and 4.5-5 mm diam., with 10 leaves, and a portion
of a large twig, 10-11 mm diam., with a single leaf and a flower borne on a
lateral twig, 4-5 mm diam.; also a single fruit. No spines are shown
on either twig.
The plate shows the following
characters: Leaves lanceolate, with acute or slightly acuminate (one leaf is
obtuse) apices, gradually narrowed, then abruptly rounded at the base; leaf
blades 9-13 X 3.5-5.5 cm, glabrous above (?) but with scattered hairs below,
veins 6-7 pairs, visible on both surfaces; petioles 9-15 mm long, 2.2-3 mm
diam., pubescent; flowers arising singly or 2 (or 3?) in the axils of the
leaves, 18-23 mm long, pedicels 6-8 mm long, 2 mm wide, with small bractlets
near the base, pubescent like the petioles; calyx 4.5-6 mm wide, 4-4.5 mm long,
with acute lobes 1.5-2.5 mm long; petals strap-shaped, narrowed toward the
rounded tips, 18-20 X 3-4 mm, glabrous; staminal filaments 13-15 mm long,
slender, glabrous, anthers short, 3 X 1.5 mm (less than 1/4 the length of the
filaments), long oval; pistil 18-21 mm long; ovary subglobose, 4.5 mm diam.,
glabrous; style glabrous, 14-17 X 1-1.1 mm, stigma capitate, 3-3.8 mm diam.;
fruits ovoid, 25-30 mm, bluntly pointed at apex, rounded at base, finely
pubescent (?), borne on a subglobose (?) gynophore (4.5 mm diam. below) which
remains attached to the apparently glabrous persistent calyx, no larger than
when the flower opened. The plate gives every evidence of having
been made carefully and accurately from a good flowering specimen.
Hooker (1875, vol. 1, p. 510) described P.
griffithii (evidently named in honor of Griffith) and included Griffith's
Citrus scandens,which he stated has very small flowers (only 6 to 8 mm
long); yet Griffith's plate shows flowers 15 to 18 mm long. Hooker
stated: "Griffith's figure is a great exaggeration, as his specimens show," but
so great an exaggeration as this seems improbable, especially since the plate
looks like a good cut made from the living plant. Possibly the best
material was used to make the plate and the dried specimens that were put in
the herbarium do not show the flowers properly and may even belong to another
species.
Griffith described his plant as a
species of Citrus, and because it was so utterly different from any known
Citrus a detailed description probably seemed unnecessary. It
would not be surprising if Griffith mixed up two species of Paramignya in
his material of Citrus scandens.
It
seems best for the present to let P. scandens stand as described and
figured by Griffith and avoid attempting to include in it the widely different
forms that have since been assigned to it. New material from the
type locality, Mogoung Valley (?) in northern Burma, is needed to settle the
status of this species.
2a.
Paramignya scandens subsp. ridleyi (Burkill) Swing. Jour. Wash.
Acad. Sci. 28:533. 1938. Paramignya ridleyi Burkill, Gard. Bul.
Straits Settl. 5:214. 1931; P. citrifolia var. beta
(pubescens) Oliv. Jour. Linn. Soc. 5(Suppl. 2):42. 1861; P.
griffithii Hook. f. 1875, in part (only as regards the Malacca specimen,
fide Ridley); P. griffithii Ridley (non Hook f.), Fl. Malay Penin.
1:356. 1922.
Type.—Singapore
(Hullett).
Distribution.—Malay
Peninsula: Malacca and Singapore.
Ridley
described the Malay Peninsula plant under the name P. griffithii, as
follows: "Shrubs shortly spiny, glabrous or pubescent. Leaves
coriaceous, elliptic or oblong, short, blunt-acuminate, 3 to 5 in. [7.5-12.5
cm] long, 1.5 to 2.3 in. [3.8-5.8 cm] wide, base rounded. Flowers
solitary or fascicled, axillary, slender, 0.3 in. [6 mm] long, white,
fragrant. Calyx acutely 5-lobed. Petals oblong, 0.3 in.
[7.5 mm] long, blunt. Stamens 10, filaments hairy, longer than the
linear-oblong anthers. Ovary 5-celled; ovules 2 in each cell."
Burkill (1931, p. 214), in making a new species
for this plant, wrote: "P. Griffithii Hook. fil in loc. cit., [Hooker f.,
1875, vol. 1, p. 510] only as regards the Malacca specimen: Ridley, Fl. Mal.
Penin., 1, 1922, p. 356. This species has much longer pedicels than
P. scandens and is abundantly distinct. It was collected in
Malacca by Griffith, and by Hullett in Singapore. The backs of the
leaves are less hairy than those of P. scandens."
Unfortunately, in the absence of type material,
no additional details can be given for this subspecies. It must
therefore, for the present at least, be kept distinct not only from P.
scandens, but from P. monophylla (= Atalantia correae Guill.),
all of which were considered by Tanaka (1930a, p. 162) to be P.
scandens.
Even as matters stand, the
subspecies is fairly well delimited by its broad elliptic-oblong leaves (7.5 to
13 by 3.8 to 6 cm) and its very small flowers borne singly or in few-flowered
axillary fascicles, with acute calyx lobes and blunt-oblong petals 6 to 7.5 mm
long, and hairy filaments longer than the anthers.
It differs from P. scandens subsp.
hispida in being glabrous or less pubescent with respect to leaves, calyx,
and gynophore; from P. andamanica in having much smaller leaves, smaller
flowers with larger calyx (?) and shorter pedicels; from the little-known P.
citrifolia in having longer pedicels and filaments longer than the anthers;
from P. armata in having much larger leaves, with longer petioles and
stouter pedicels. It differs from P. surasiana and P.
rectispinosa by having styles only three times as long as the ovary.
2b. Paramignya scandens
subsp. hispida (Pierre) Swing. Jour. Wash. Acad. Sci. 28:533.
1938. Atalantia hispida Pierre ex Guill. in Lecomte, Not. Syst.
1:182. 1910.
Type.—French
Indo-China, Cambodia, Bienhoa Province.
Distribution.—Known only from the type locality.
Differs from the species in having longer
petioles and smaller flowers with shorter, broader petals. It
resembles more closely the subspecies ridleyi but has somewhat smaller
leaves and anthers apparently shorter in proportion to the length of the
filaments, and possibly longer recurved spines.
Guillaumin (1911, p. 674) described Atalantia
hispida Pierre about as follows: Branches slender, at first very pubescent,
later on becoming glabrous, with spines about 10 mm long; leaves oval or
oval-oblong, 6-10 X 2-4 cm, with a short acuminate tip, rounded or subcordate
at the base, velvety-pubescent below, veins invisible on upper surface,
parallel and close together below; petiole velvety-pubescent, up to more than 2
cm in length; inflorescences solitary, axillary; flowers 10 mm long, on velvety
pubescent pedicels 5 mm long, with small bracts at the base; calyx cup-shaped,
2-3 mm long, sepals 5, oblong, acute (valvate in flower bud), pubescent
without, glabrous within; petals 5, elliptical, 1 cm long, glandular, slightly
pubescent without; stamens 10, 2/3 length of petals, filaments dilated,
pubescent, usually free (often 2 or 3 connate), anthers elliptical, half as
long as the filaments, not apiculate; gynophore cylindric, pubescent,
glandular, crenulate above; ovary ovoid-globular, 5-angled, glabrous, shorter
than the style, with 5 locules, each containing 2 ovules, style cylindrical,
glabrous, stigma capitate, slightly 5-lobed; fruits small (according to
Pierre).
Tanaka (1928b, p. 712)
considered this subspecies to be a synonym of P. scandens (Griff.) Craib,
but it appears to have smaller flowers with decidedly shorter and broader
petals. It will not be possible to understand clearly these three
forms here referred to P. scandens until studies can be made on
collections of all three in comparable states.
3. Paramignya armata (Thwaites) Oliv. Jour. Linn. Soc. Bot.
5(2):43. 1861. Arthromischus armatus Thwaites, Enum. Pl. Zeyl. 47.
1858.
Type.—Ceylon (Thwaites, No.
1187). Herb. Kew.
Distribution.—Ceylon (hotter parts of the island).
Leaves oval, elliptic or elliptic-lanceolate,
obtusely caudate-acuminate, rounded at the base, 5-8 X 2-3.5 cm; petioles short
(8-11 mm); pulvinus 3 mm long, articulated with the petiole and with the leaf
blade; flowers small (6 mm long), solitary or in few-flowered axillary
clusters, 4-5-merous; pedicels very slender (0.4-0.5 mm diam.), 6-8 mm long;
calyx minute, obtusely 4-5-lobed, lobes acute; petals oblong, very much
imbricated; filaments linear-subulate, pubescent, slightly dilated in the
middle; ovary pubescent, borne on a gynophore equaling it in height, with 3-4
locules, ovules usually 1 in each locule; style long, glabrous, cylindric,
stigma globose; fruit subglobose, apiculate, 10-20 mm diam., yellow, 1-4
seeded.
This species is said to have pulpy
fruits but nothing is known about the nature of this pulp.
Thwaites directed attention to its "jointed leaf
stalk," a very striking character often overlooked by subsequent
authors. The petiole shows a stiff straight petiole proper, at the
tip of which is articulated a short fleshy pulvinus.
4. Paramignya beddomei Tan.
Jour. Bot. Brit. & For. 68:230. 1930. Paramignya armata
Beddome (non Oliver), Icon. Pl. Ind. Or. 65, pl. 275 (excl. descript.). 1874.
Illus. Beddome, loc. cit. pl. 275 (as P.
armata).
Type.—British India,
Madras, Anamaly (Anaimalai Hills) (Beddome, No. 1052). Herb. Brit.
Mus., London.
Distribution.—South
peninsular India and Ceylon (fide Tanaka).
Twigs very slender, conspicuously zigzag,
pubescent with fine appressed hairs; leaves medium-sized, blade 70-75 X 35-40
mm, ovate, caudate, broadly rounded at base, pubescent on the midrib,
especially below, petiole long (10-15 mm), sigmoid with an articulated pulvinus
occupying the upper third, pubescent; flower buds large (10-12 X 3-4 mm),
cylindric; flowers solitary, axillary, large (25-30 mm diam. when open)
pedicels 8-12 mm long; calyx cupulate, lobes short, obtuse, auriculate; petals
linear, bluntly rounded, 15 X 4-5 mm; filaments apiculate, very pubescent for
their whole length, anthers linear, but contorted at maturity; ovary 4-5-lobed,
cylindric, almost glabrous, but sparingly pubescent in upper portion, about as
long as the disk, ovules 1 in each locule; style linear, very glandular, stigma
depressed-globose, 5-lobed; fruit globose, not apiculate.
Beddome's figure (l.c., pl. 275) shows short
recurved single spines, 6 to 12 mm long, tapering to a fine point.
This species has been confused by previous students of Paramignya with
P. armata, from which Tanaka separates it by many characters.
5. Paramignya citrifolia
(Roxb.) Oliv. Jour. Linn. Soc. Bot. 5(2):42 (exel. descript.; valid for
name only). 1861. Limonia citrifolia Roxb. Fl. Ind. 2:379. 1832;
Paramignya citrifolia Hook. f. Fl. Brit. Ind. 1:510 (excl. synon.;
valid for descript. only). 1875; Atalantia citrifolia (Roxb.) Kurz, Jour.
Asiat. Soc. Bengal 39(2):69. 1870.
Type.—Northeastern British India near Burmese boundary, Chittagong
(Roxburgh, No. 340). Herb. Brit. Mus.,
London.
Distribution.—Reported from
Bengal and northeastern India (now partly East Pakistan) by Tanaka
(1930b, p. 61).
Roxburgh's original
description (under the genus Limonia) reads: "Shrubby; armed with
recurved spines. Leaves simple, elliptically oval, entire,
obtusely acuminate. Flowers axillary.
Berries ovate, few-seeded.
"A very
ramous, rigid, well-armed shrub, of five or six feet in height, a native of the
forests of Chittagong, and with the other armed species, well adapted for
fences. Flowering time the hot season.
"Young shoots polished.
Thorns axillary, solitary, short, somewhat recurved.
Leaves alternate, round-petioled, elliptic, with an obtuse, somewhat
lengthened point, entire, smooth, but marked with numerous pellucid points, as
in many Aurantiae; from four to five inches long, and from two to three
broad. Stipules none. Flowers small,
white, short-peduncled, axillary. Bracts minute, about the
insertion of the peduncles, and on them. Calyx five-toothed,
having its substance marked with pellucid points. Petals
five, oblong, smooth. Filaments ten, distinct, short,
inserted round the base of the germ. Anthers linear,
erect. Germ [ovary] ovate-oblong, five-grooved, on the
outside five-celled, each cell containing two ovula attached to the
axis. Style thick and short. Stigma
sub-peltate. Berry ovate, of the colour and appearance of a
lime, even to the little green cells in the cortex. Seeds
from one to four, separated by some few small fibers only, which are
scarcely to be traced when dry, oblong, having the sides agreeing in shape
with the number in the berry. Integument single,
membranaceous. Perisperm none. Embryo
inverse. Cotyledons conform to the seed.
Radicle superior."
The type specimen in
the British Museum (Roxburgh, No. 340; see Tanaka, 1930b) of two fruiting
twigs from Korelea Hill, collected in 1811 (?), shows twigs, slender ultimate
growth, 2-3 mm diam.; leaves 6-12 X 3-6 cm, elliptical, abruptly acuminate but
blunt at tip, base gradually rounded, seldom very broadly cuneate, 12-14 pairs
of lateral veins, inconspicuous above but visible on under side, arising at a
wide angle (65°-70°) with the midrib; petioles 8-10 mm long, not clearly
jointed to the pulvinus; fruit subglobose, 12 X 14 mm, pedicel 6-8 X 1.5 mm, on
which the calyx, 5 mm wide but only 2-3 mm long, persists. These
notes are based on a photograph by Tanaka (No. 3789).
Hooker, who compiled his description (without
seeing any specimens) from Roxburgh's description and figure (Mus. [E.I.C.]
2243, fide Oliver, 1861), found the following characters: "…Young shoots
polished. Leaflet 4-5 by 2-3 in., quite entire.
Flowers white, about 1/3 in. diam. Petals
oblong. Filaments quite free. Ovary
ovoid-oblong, 5-grooved; style short, thick; stigma subpeltate.
Berry ovoid, pointed, of the colour and appearance of a lime, even
to the little green cells in the cortex. Seeds 1-4, separated
by a few small dry fibers only, which are scarcely to be traced when dry,
oblong; testa membranous.—This plant is unknown to me; the description is taken
from Roxburgh's Flora Indica and drawings. Prof. Oliver
referred to it in the preceding species [P. griffithii], which differs in
the form of the stamens, and (if Roxburgh's figure of the ovary is correct) in
wanting a style, but the said figures resemble what an ovary would be were the
style fallen away. I refer it to Paramignya from the long
anther and pointed fruit. Kurz has pointed out that Oliver's P.
citrifolia and Griffith's Citrus scandens cannot be the same as
Roxburgh's Limonia citrifolia (Jour. Asiat. Soc. Bengal, 1870,
Pt.2, 69)."
This very imperfectly known
species has been found with certainty only near Chittagong not far from the
frontier between East Pakistan and northwestern Burma. It is placed
by Hooker in the group of species having "flowers about 1/2 inch [12-13 mm]
long. Calyx small with acute teeth," in which group he also places
P. griffithii and P. armata.
It
would be very desirable to secure more good herbarium specimens from the type
locality, Chittagong. Until this is done, it will be very
difficult, if not impossible, to determine with any certitude the affinities of
P. citrifolia.
6.
Paramignya grandiflora Oliv. Jour. Linn. Soc. Bot. 5(2):42.
1861.
Type.—Southern Burma, Tenasserim
(?) (Lobb, No. 925). Herb. Kew.
Distribution.—Southern Burma: Tenasserim; Andaman Islands; Malay Peninsula
(?).
Much like P. monophylla, having
equally large, solitary flowers, 18-19 mm long, but broader leaves, broadly
rounded at the base, pubescent below; petiole articulated with the pulvinus
(?); calyx cupular, lobes well developed, often cuneate at tips and overlapping
(imbricate?) at the base, ciliate; fruits unknown.
This species is native in southern Burma and the
Malay Peninsula and is much like P. monophylla, having large flowers with
five petals; the styles pubescent below and the stamens pubescent and similar in
shape to those of P. monophylla. The type collection was
credited to Singapore by Oliver (1861, p. 42) and is so labeled, but, according
to Ridley (1922, p. 355), erroneously. He suggested that it is from
Tenasserim and stated: "it is otherwise known only from Tenasserim" (where it
was collected by Wallich, No. 6361).
7. Paramignya andamanica (King) Tan. Bul. Soc. Bot. France
75:712. 1928. Paramignya armata var. andamanica King, Jour.
Asiat. Soc. Bengal 62(2):223. 1894.
Type.—Andaman Islands (King). Herb.
Kew.
Distribution.—Andaman Islands,
North Vietnam, South Vietnam, Sumatra, Sumbawa, Sabah (North Borneo.)
King's original description and his type
specimen in the herbarium at Kew give the following characters: Twigs slender,
puberulous when young, soon becoming glabrous; spines single, short, curved,
pubescent; leaves large, elliptical or elliptic-oblong, narrowed at apex into a
blunt point, broadly rounded at base, 10-23 X 3.5-9 cm, with 10-12 pairs of
veins making an angle of 65°-75° with the midrib; petiole 12-25 mm, pubescent
on the upper surface when young, not distinctly articulated with the pulvinus;
flowers 5-merous, paired or solitary, axillary; flower buds slender, 14-18 mm
long; pedicels long, slender (18-20 mm), arising separately; calyx glabrous,
short (3-4 mm), deeply divided into 5 broadly obtuse lobes; petals about 4-5
times as long as the calyx (15-16 mm), linear-oblong, obtuse; stamens free,
almost as long as the petals, filaments rather thick and woolly for the lower
three-fourths but filiform and glabrous above; ovary glabrous, ovoid, seated on
a short cylindric disk not broader than the ovary; style elongate, slender,
glabrous; fruits globular or turbinate, glabrous, 12-16 mm diam.
Parkinson (1923, p. 108) reported it as "fairly
frequent" in several of the Andaman Islands. His specimens were
more vigorous than those seen by King, as he described it as having curved
spines about 5 cm long, leaves 10-23 X 4-9 cm, with petioles 25 mm long (all of
these measurements are the largest yet reported for any species of
Paramignya); flowers small (12-13 mm long), on slender pedicels 25-38 mm
long; filaments pubescent; fruits 12-13 mm diam., globular or obscurely
2-lobed. He found the leaves "almost scentless when bruised."
Guillaumin (1911, p. 675) reported this species,
under the name P. aramata, as occurring in Cochin China (South
Vietnam). He described it as having curved spines only 6-12 mm
long; leaves 7.5-12 X 3.5-4.5 cm; flowers on slender pedicels 30 mm long; calyx
short (3 mm), lobes 5, indistinct, rounded, ciliate; petals 3 times as long as
the calyx; stamens almost as long as the petals, dilated and pubescent at the
base, slender and glabrous at the tips; ovary globose, glabrous, 5-sided, with
a style 3 times as long; stigma capitate, flattened above, attaining height of
the stamens; fruit yellow, globose, borne on a pedicel 30-35 mm long, showing
the remains of the eccentrically placed style.
This remarkable but little-known plant reaches
its best development in the Andaman Islands, although it grows also in South
Vietnam, and, according to Tanaka (1931b, p. 10), in Sumatra, Borneo,
and Sumbawa, which lie about 1,000 to 3,000 kilometers to the
southeast. The Andamans are situated about 1,300 kilometers nearly
west from South Vietnam.
It is curious that
this species has not been reported from southern Burma, southern Thailand, and
the Malay Peninsula. Possibly some of the small-flowered forms of
Paramignya reported from the Malay Peninsula belong to this species as
varieties or subspecies rather than to P. scandens or P.
citrifolia.
This species, like
Atalantia macrophylla (which also is native to the Andaman Islands), is
remarkable for its extraordinary vigor, especially in its native habitat.
It is the only species of Paramignya that
approaches the dimensions of some of the species of Luvunga (the other
genus of woody vines belonging to the orange subfamily) in the length of its
leaves and spines.
This species is related to
P. armata, of Ceylon, in having very similar small flowers, but differs
in having much larger leaves and spines, and, as King stated, in having fewer
flowers in the leaf axils.
8.
Paramignya longipedunculata Merr. Phil. Gov. Lab. Bur. Bul. 35:24.
1905. Atalantia longipedunculata (Merr.) Guill. in Lecomte, Not. Syst.
1:221. 1910.
Type.—Philippines,
Luzon Island, Rizal Province, Bosoboso (Ahern's collector). Herb.
Bur. Sci., Manila.
Distribution.—Philippines: Luzon and Mindoro
islands.
Common name.—Luzon
paramignya.
Branches hairy, with spines 5-15
mm long, retrorse and slightly curved, densely pubescent when young, but the
brownish, hardened tips glabrescent with age; terminal twigs dark green when
dry, densely pubescent; leaves oval or elliptic-ovate, 7-10 X 5-7 cm, with the
apex abruptly acute or broadly short-acuminate, base rounded, pubescent on
lower surface, midrib densely pubescent below; petioles 8-10 mm long, densely
pubescent, pulvinus not visibly articulated with the leaf blade or petiole;
flowers solitary or in 2- or 3-flowered axillary clusters; pedicels densely
pubescent, 1.5-3 cm long, subtended by slender pubescent bracts, 1.1-2 mm long,
borne at the tips of short branches, 2.5-7 mm long; calyx cupular, densely
pubescent, 7 mm diam., 5-lobed, lobes erect, broad, ovate, 2.5 mm long; petals
5, linear-oblong, 20 X 5 mm, densely pubescent without: stamens 10, filaments
pubescent, 12 mm long, anthers 5 mm long; ovary with 5 locules; style about 15
mm long, densely pubescent; fruits broadly ellipsoid, bluntly pointed at apex,
conoid at base, 2.5-3 mm X 1.5-1.8 cm, subtended by the persistent calyx.
This is a large-flowered species like P.
monophylla, but differs in being more or less densely pubescent in all
parts. It also has much longer pedicels than that
species. Merrill (1923, p. 339) reported that it grows "in
forested ravines and in secondary forests at low and medium altitudes."
9. Paramignya mindanaensis
Merr. Phil. Jour. Sci. Bot. 3:140.
1908.
Type.—Philippines, Mindanao
Island, Lake Lanao, Camp Keithley (Mary Strong Clemens). Herb. Bur.
Sci., Manila.
Distribution.—Philippines: Mindanao Island; also Samar and Luzon islands
(fide Tanaka).
Branches glabrous except
the flowering twigs; leaves oblong-elliptical, glabrous, 9-12 X 3-6 cm, apex
acuminate, base rounded, veins indistinct; petioles 1 cm long; flowers, 1 or 2,
axillary, about 17 mm long; pedicels slender, 10-15 mm long; calyx cupular,
slightly hairy, about 5 mm diam., 5-lobed, the lobes 1.5 mm long, rounded;
petals imbricate, oblong, 12 X 5 mm, glabrous; stamens 10, filaments thickened,
somewhat pubescent, about 7 mm long, anthers 4 mm long; gynophore about 2 mm
long and 2 mm diam., crenulate; pistil 10 mm long, somewhat pubescent; ovary
5-angled, with 5 locules; style stout; fruits (immature) 1.5-2 cm diam.,
usually curved, glabrous.
This species differs
from P. longipedunculata of Luzon Island in being, on the whole, less
pubescent, since some parts are glabrous, and in having larger leaves and
smaller flowers. It has been referred to P. grandiflora as a
synonym by Tanaka (1932e, p. 425), who stated that the type specimen of
P. mindanaensis is a typical P. grandiflora. However,
one of Lobb's specimens of the latter species from Singapore, cited by Oliver
in the original description, shows pedicels measuring 20 to 26 mm long, instead
of 10 to 15 mm, as in Merrill's description of P. mindanaensis, and
calyxes nearly twice as large as in the Mindanao species. Inasmuch
as P. mindanaensis occurs 1,500 miles east of Singapore, it would seem
best to maintain it as a good species for the present. Tanaka found
that specimens from Luzon and Samar islands are much less pubescent than the
type material from Mindanao Island.
10. Paramignya surasiana Craib, Kew Bul. Misc. Inform.
1916:261. 1916. Paramignya griffithii Hook. f. Fl. Brit. Ind.
1:510 (pro parte) (so far as concerns the specimen from Pegu collected by
McClelland, fide Craib). 1875.
Type.—Thailand, Chiangmai, Doi Sutep (Kerr, No.
2542).
Distribution.—Thailand; northern
Burma: Pegu.
[After Craib]: Twigs puberulous;
spines recurved, 7-9 mm long; leaves obovate or elliptic-obovate, apex obtusely
caudate-acuminate, base broadly cuneate or rounded, 10-14 cm long (including
the acumen, which may be up to 1 cm long), 5.2-7.5 cm wide, chartaceous,
glabrous except for the shortly pubescent midrib, oil glands abundant,
conspicuous on the lower surface, lateral veins straight, at least 10 on each
side, prominent on the underside; petiole 1.5-1.7 cm long, puberulous; pedicels
about 4 mm long with a few bractlets below; flower buds oblong, rounded at the
apex, up to 1.4 cm long; calyx 5.5 mm long on the outside, curly pubescent
without (like the pedicels), appressed pubescent within, lobes unequal in
length, 1-2.5 mm long, with rounded ciliate tips; petals white, at least 1.7 cm
long, glabrous; filaments 12 mm long, angular, flattened at the middle, nearly
glabrous at base, elsewhere white pubescent; ovaries short, densely pubescent;
style about 9 mm long, pubescent.
This species
is still inadequately known. The fruits, said by Craib to have been
collected by McClelland at Pegu in Burma, are not described by either J. D.
Hooker or Craib. The petals are very imperfectly known, and no
measurements of the ovary are available. However, since Craib
described this species along with P. rectispinosa, which has an
exceptionally short ovary, and furthermore described the ovary of P.
surasiana as "short," it is clear that this species also has a very short
ovary. Even if the ovary of P. surasiana were twice as long
as that of P. rectispinosa, it would still be less than one-fourth as
long as the style. More and better material of this species is much
needed in order to place it accurately. It was collected by Kerr
in evergreen jungle in the same locality in which he discovered P.
rectispinosa but at an altitude nearly 1,000 m higher, i.e., at 1,650 m
instead of 660 m.
11.
Paramignya confertifolia Swing. Jour. Arnold Arbor. 21:17. 1940.
Illus. Swing. loc. cit. pl. 4, figs. 1, 2; fig. 3-17 this work.
Type.—China, Hainan, San Tsuen Mountain (Tsang Wai Tak, No.
15523). Herb. Univ. Calif.,
Berkeley.
Distribution.—Southeastern
China: Hainan Island and Kwangtung Province.
A
shrub, 3-5 m high, clambering over shrubs; young twigs slender, terete,
pubescent, with internodes 2-5 cm long on vigorous shoots but much shorter,
often only 8-10 mm long, on fruiting branches; spines short (3-10 mm; usually
3-6 mm), recurved, often reduced in size or wanting on the end branches,
especially on fruiting branches; leaves oval or oblong to long elliptical,
usually broadly rounded at the base (sometimes cuneate on longer leaves), the
very tip of the acumen bluntly rounded, midrib and lateral veins visible on
both surfaces but more distinct below, the 8-12 or more pairs of lateral veins
arising at an angle of 60°-70° with the midrib, the space between being filled
with smaller more or less parallel veinlets making elongate reticulations,
margins entire or irregularly and shallowly crenulate; petioles 4-12 X 1-1.5
mm, flattened above, more or less pubescent, wrinkled in dried specimens,
pulvinoid portion not articulated, the whole petiole curving more or less to
place the leaf blades in best position to catch the incident light; flowers
axillary, arising singly or in small clusters, sometimes in greatly reduced
racemes; flower buds cylindrical, 8-10 X 2.5-3.5 mm; pedicels slender, 4-5 mm
long, 0.5-0.7 mm wide, glabrous, subtended by minute sparsely hirsute bracts
where the pedicels join the peduncles, which are hirsute; calyx small, 2 mm
wide, 1-5 mm high, brownish buff-colored (as is the pedicel), lobes 5,
triangular, with ciliate tips; petals 5, glabrous, white when fresh,
yellowish-brown when dry, 7-9 or 10 X 3-4 mm; stamens 10, filaments flattened,
5-6 mm long, anthers linear, 1.8-2 mm long; disk cylindrical, not broader than
the ovary base; ovary ovoid, 1.2-1.3 X 1 mm, strongly hirsute with
yellowish-buff hairs, with 2 locules, each with 2 ovules, narrowed abruptly
into the sparsely hirsute style, 4.5-6 X 0.3-0.4 mm wide at base and more
slender above, dehiscent shortly after flowers open, stigma depressed globose,
0.5 mm high and 1-1.5 mm wide; fruits at first sub-globose, glabrous, but when
full-sized becoming very rough, sometimes almost lobed with irregularly rugose
folds of the peel, which shows numerous sunken oil glands having a
turpentine-like odor and flavor, yellow when ripe, subglobose, 1.5-2 cm diam.,
apparently corky in places; seeds small, ovoid, monoembryonic.
This species has been identified as P.
scandens (Griff.) Craib, and as P. griffithii Hook. f., but it seems
rather to be a new species allied to P. surasiana Craib from Thailand in
having a very small ovary only a fraction of the size of that of the species
found to the west of Thailand. It differs from P. surasiana
in a number of important characters and seems to be a good new species (see
fig.
3-17). The ovary of P. confertifolia is slightly less
than a fourth as long as the style (see Swingle, 1940a, pl. 4, fig.
2).
12. Paramignya
rectispinosa Craib, Kew Bul. Misc. Inform. 1916:261. 1916.
Atalantia griffithii Craib (non Guill.), Aberdeen Univ. Studies
57:53. 1915 (?).
Type.—Thailand,
Chiangmai, Doi Sutep (Kerr No. 1718). Herb.
Kew.
Distribution.—Thailand.
[After Craib]: A climbing shrub with green,
fleshy pubescent (soon puberulent twigs) [sic], up to 3.5 mm diam., with
axillary, straight or nearly straight, pubescent spines nearly 2 cm long, green
except for the straw-colored tips; leaves oblong or oblong-oblanceolate, rigid,
chartaceous, obtusely-acuminate at the apex, gradually narrowed toward the
rounded base, 10.5-14 X 2.8-4.7 cm, blades glabrous above except for the
midrib, which is finely pubescent (very rarely subglabrous) especially at base,
soft-pubescent below, with many pellucid dots [oil glands], lateral veins about
15 on each side, anastomosing inconspicuously, faint above but prominent below;
petioles 10-11 mm long, slightly channeled above, pubescent like the twigs;
flowers white (fide Kerr), solitary; pedicels about 3 mm long,
pubescent, with small branchlets near the base; calyx 2 mm long, pubescent
without, glabrous within, 5-lobed, lobes deltoid or narrow-deltoid, rather
obtuse, slightly shorter than the tube; petals oblong, 12 X 3 mm, glabrous,
conspicuously glandular dotted; filaments 8 mm long, pubescent, glandular like
the petals, anthers 2 mm long; disk slightly shorter than the calyx; ovary
about 1 mm long, densely pubescent; style 7 mm long, with divergent hairs;
stigma capitate.
Craib compared this species
with P. griffithii Hook. f. [= P. scandens], "from which it
differs in having spines nearly 2 cm long, straight or nearly
straight." This species is known only from Craib's original
description, which shows it to differ markedly from the other species of the
genus, not only in the long, straight (or nearly straight) spines, but also in
its short calyx, very short ovary, and short anthers. The type
specimen was collected in evergreen jungle at 660 m above sea
level.
SPECIES OF DOUBTFUL RELATIONSHIPS
13. Paramignya cuspidata (Ridl.) Swing. Jour. Wash. Acad.
Sci. 28:533. 1938. Atalantia cuspidata Ridl. Jour. Straits Branch
Roy. Asiat. Soc. 82:174. 1920.
Types.—Malacca, Sungei Hudang (Ridley, No. 8391), and Dindings, Lumut
(Ridley, No. 7944). Herb. Hort. Bot.,
Singapore.
Distribution.—Known only
from the type localities in the Malay Peninsula.
The original description reads as follows: "A
spiny bush; branchlets pubescent; spines short, blunt, straight.
Leaves thin-textured, not coriaceous, ovate-elliptic, cuspidate, blunt, base
round, pubescent on the back, midrib sunk above, elevated beneath, the nerves
about 6 pairs, very fine, forked at tip, 2-3 in. [5-7.6 cm] long, 0.5 in. [12
mm] wide; petiole 0.2 in. [5 mm] long, pubescent, jointed in the
middle. Flowers solitary; pedicels 0.25 in, [6 mm]
long. Sepals round, pubescent. Petals 5, linear,
oblong, 0.4 in. [1 cm] long, white. Stamens linear,
oblong. Ovary cylindric, hairy. Style stout,
glabrous. Stigma orbicular. Fruit ovoid, blunt, 0.8
in. [2 cm] long; rind thin, pulp scanty; seeds 2, large flattened."
Thanks to the kindness of I. Henry Burkill,
former director of the Singapore Botanic Gardens, Swingle secured a photograph
from Kew Gardens of one of the type specimens collected by H. N. Ridley (No.
7944) at Lumut, Dindings, March, 1890. A close study of this
photograph shows that the petiole is twisted just as those of other species of
Paramignya, because of a pulvinus just below the leaf blade, noted by
Ridley who stated: "petioles…jointed in the middle." This refers to
the junction of the pulvinus with the lower rigid portion of the
petiole. As the plant does not have recurved spines like true
species of Paramignya, it would seem at first sight not to belong in this
genus. However, an inspection shows that P. cuspidata has
what seem to be leafy side branches that arise at right angles and sometimes
curve backward. The pulvinate pubescent petioles, the recurved side
branches, and the general character of the leaves seem to indicate either an
ancestral form from which the true Paramignya have evolved or else a
degenerate form of Paramignya that has largely lost its climbing habit
and no longer has recurved spines. This species is connected with
the group of normal long-spined species of Paramignya through P.
lobata, which is a woody climber with short reverted spines.
14. Paramignya lobata Burkill,
Gard. Bul. Straits Settl. 5:214. 1931. Atalantia hispida Ridl.
(non Pierre), Fl. Malay Penin. 1:357. 1922; A. monophylla Ridl.
(non DC.), Jour. Straits Branch Roy. Asiat. Soc. 26:26.
1894.
Type.—Johore, Tumpat near Kota
Pahat (Lake and Kelsall. Nov. 11, 1892). Herb. Hort. Bot.,
Singapore. Cotype: Kelantan, Tumpat near Kota Bakru (Ridley,
Feb., 1917). Herb. Hort. Bot.,
Singapore.
Distribution.—Malay
Peninsula.
A climbing vine, with short (4-8
mm), retrorsely curved, stout, sharp spines with strong bases oval in cross
section; twigs not angled but with slight longitudinal ridges, glabrous,
apparently also slightly glaucous, internodes 1-3 cm long; leaves thin but more
or less coriaceous, oblong-oval or long-elliptical, gradually acuminate, the
extreme apex of the acumen bluntly rounded or subacute, bases broadly rounded
or even slightly cordate, 6-12 X 2.2-5 cm, margins subentire, slightly undulate,
surface densely and evenly dotted with oil glands, midrib depressed above,
strongly salient below, lateral veins faint above, and not strongly marked
below, 12-15 pairs arising from the midrib at an angle of 55°-60°, connected by
coarsely reticulate veinlets; petioles 5-8 X 1-2 mm, wingless, but with a narrow
furrow on the upper surface, glabrous, not clearly articulated with the leaf
blade and not jointed in the middle, the entire petiole more or less pulvinoid
and able to bend and twist to put the leaf blade in proper position to catch
incident light; flowers solitary, axillary; sepals 5, oblong-acute, becoming
triangular, ciliate or velvety-pubescent without; petals 5, elliptic, slightly
pubescent without, 7-8 mm long; stamens 10, filaments free, velvety-pubescent;
disk a cylindric gynophore, velvety-pubescent; fruits red, thin-walled, 1-1.5
cm long, 1-2 cm diam., deeply 2-5-lobed, depending on the number of seeds
formed; pedicels 6-8 X 0.7-1.5 mm, more or less longitudinally ridged in the
dried specimens; seeds about 9 X 6 X 4 mm, apparently monoembryonic.
Thanks to the courtesy of R. E. Holttum, then
director of the Botanic Gardens, Singapore, who sent two type specimens on
which this species was based, Swingle had the best possible material to study
except for the flowers, which are known only from Ridley's description of this
plant cited above. The strikingly lobed fruits of this species are
quite unlike those of any other species of Paramignya, and are for that
matter unique in the tribe Citreae, although they are somewhat similar to the
lobed fruits of Murraya gleniei of the tribe
Clauseneae.
Paramignya lobata resembles
the shrubby P. cuspidata in having short, stout spines. Both
of these species need further detailed study to determine more definitely their
true affinities.
15.
Paramignya trimera (Oliv.) Burkill, Gard. Bul. Straits Settl.
5:213. 1931. Atalantia trimera Oliv. Jour. Linn. Soc. Bot.
5(2):24. 1861; Triphasia monophylla DC. 1824; Atalantia (?)
recurva Benth. 1863.
Type.—East
Indian Archipelago, Timor Island (L'Eschenault, Tanaka's Det. No.
Q-1719). Herb. Kew.
Distribution.—Java; southern Philippines: Mindoro and Mindanao islands;
Timor; northeastern Australia.
A very full
description of this species based on material from Timor (the type locality)
published by Decaisne (1834, v. 439) under the name Triphasia monophylla
D. C., reads in translation as follows: "Twigs erect, rounded, bark
glabrous, verrucose, yellow-gray when young, becoming green. Leaves
simple, 1-2 in. [2.5-5 cm] long, 3/4 in. [18 mm] wide, oblong-ovate,
subentire, sometimes emarginate at the apex, midrib prominent below,
coriaceous, shiny, punctate with numerous pellucid glands, petiolate; petiole
4 -2 lin. [8-4 mm] long, somewhat glabrous. Spinescent stipules
[spines] acute, 4-6 lin. [8-12 mm] long, horizontal, with a greenish cortex,
becoming gray. Inflorescence racemose, axillary, half as long as
the leaves, 8-2 flowers; pedicels very short, somewhat glabrous, with 3
bracts. Calyx persistent, 3-toothed, glandular-punctate, with
subrounded teeth, concave, margin ciliolate. Petals 3, rarely 4,
alternating with the calyx, 2 lin. [4 mm] long, obovate, concave,
subcoriaceous, without veins, glandular, punctate in the middle,
smooth. Stamens 6, shorter than the petals, filaments thick,
flattened at the base, free, without glands, equaling the ovary in
length. Anthers oblong-sagitate, obtuse at the apex, subglandular,
punctate, longitudinally dehiscent. Pistil almost equal to the
stamens in length. Ovary borne on a subsinuate disk, more or less
cone-shaped, 2-locular, 1 pendent ovule in each locule. Style
thick, glandular-punctate like the ovary. Stigma flattish,
obscurely 3-lobed. Fruit (immature) fleshy, obovoid, crowned at the
tip with the persistent stigma, filled with gum, 2-locular, 1 seed in each
locule, seeds attached at the internal angle of the segments."
A specimen collected on Mataja Island, Mindanao,
Philippines, by Kienholz (Bur. Sci., Manila, No. 15525, in Herb. Univ. Calif.),
was assigned to this species by Tanaka (1932e, p. 427). It
has leaves 8-10 X 5-6 cm, oval in outline, broadly rounded at the apex, and
rather abruptly emarginate; the wingless petioles, 5-7 mm long, are not
articulated with the blade, wrinkled when dry, doubtless pulvinoid in the fresh
state. At each node is found a single short sharp recurved spine,
5-7 mm long. The young ovaries are stipitate at the base, ovoid in
outline, tapering into a very short style, less than 1/4 the length of the
ovary, which is 2 mm long and 1 mm wide. The leaves are much larger
than those reported by Decaisne from Timor.
This species of Paramignya is an anomalous one and may prove not to
belong to this genus. It has, however, recurved spines and a
cylindrical gynophore like Paramignya. The locule walls seems
[sic] to have minute hair-like secreting emergenzen unlike any other
member of this genus yet studied.
Subtribe 2. Citrinae: Citrus
Fruit Tress
The thirteen genera of the
subtribe Citrinae (the Citrus Fruit Trees), which include sixty-five species,
differ strikingly from the other members of the orange subfamily in having
pulp-vesicles, very peculiar structures that arise from the locule walls
(especially the dorsal wall) and grow into the locular cavity, developing into
sacks filled with numerous large, very thin-walled cells full of a watery
juice. No such structures are found in any other plants of the
family Rutaceae or in related families, and no close homologies are known among
any of the higher plants. Other genera of the subfamily
Aurantioideae have secretory glands on the locule walls, from which arises the
mucilaginous gum that fills the locular cavity of the ripe fruit in most of the
genera.
The taxonomic work that has been done
since the beginning of this century on the genera most closely related to
Citrus has shown that the pulp-vesicles not only are of primary
importance in the classification of the very close relatives of Citrus
(the True Citrus Fruit Trees)7 but are even more important in
the taxonomic study of the genera included in the Near-Citrus Fruit Trees and
in the Primitive Citrus Fruit Trees. The presence or absence of
pulp-vesicles is indeed of paramount importance in the correct classification
of the small-fruited Citrus relatives and has permitted making a sharp
distinction between the two groups of small-fruited members of the subtribe
Citrinae and the deceptively similar Minor Citroid Fruit Trees belonging to the
subtribe Triphasiinae.
The thirteen genera of
the subtribe Citrinae fall naturally into three groups, A, B, and
C. Group A, which we may call the Primitive Citrus Fruit Trees,
comprises five genera, Severinia, Pleiospermium,
Burkillanthus, Limnocitrus, and Hesperethusa.
Pleiospermium has pulp-vesicles that are small, ovoid or cylindrical,
blunt-tipped, and usually distinctly corticated. Limnocitrus
has pulp-vesicles that are slender, fusiform, with very acuminate tips, somewhat
contracted at the base where they are attached to the dorsal wall of the
locule. Severinia and Hesperethusa have peripheral
pulp-vesicles that are irregular in shape and size and little differentiated in
structure. Finally, Burkillanthus, which is a very strange
genus, has pulp-vesicles of a type not found as yet in any other plant in the
orange subfamily. The walls of the pulp-vesicles are rather thick
and semitranslucent. As the fruit ripens, the pulp-vesicles
apparently collapse entirely and may perhaps set free a mucilaginous
fluid. Burkillanthus has leaf, spine, and flower characters
greatly resembling those found in the genus Pleiospermium, from which it
differs chiefly in having much larger fruits with a tough rind and in having
very numerous ovules (22 to 26) in each of the five locules, instead of two
ovules as in Pleiospermium. Probably Burkillanthus
arose from a Pleiospermium-like ancestral form.
Group B, the Near-Citrus Fruit Trees, comprises
two genera, Citropsis and Atalantia, both showing well-organized
pulp-vesicles with broad sessile bases and slender conical sides tapering to a
rather acute apex pointed toward the center of the fruit.
Group C, the True Citrus Fruit Trees, includes
six genera with highly organized pulp-vesicles that show a slender stalk at the
base (except Clymenia) and a more or less expanded fusiform body with the
apex more or less acutely pointed. The genus Clymenia has
short, often subglobose pulp-vesicles with short, usually very broad stalks or
almost sessile. Nevertheless, in all its other characters,
Clymenia is a True Citrus Fruit Tree, greatly resembling Citrus in
external character, with simple leaves like those of Citrus medica but
with venation somewhat like that in leaves of Wenzelia and
Monanthocitrus of the subtribe Triphasiinae.
The genus Hesperethusa in group A
resembles Citropsis in group B in its leaf and seed characters but has
much smaller fruits with much less definitely organized
pulp-vesicles. The genus Atalantia of group B resembles the
genera Fortunella and Eremocitrus of group C in having small
fruits with few locules, each containing only one or two ovules.
All the other genera in group C have larger fruits, with four or more (usually
six or eight) ovules in each locule.
The
twelve genera that, with Citrus, constitute the subtribe Citrinae are of
extraordinary interest since all of them are undoubtedly rather closely related
to Citrus. Some of them throw light on the origin and
evolutionary development of the True Citrus Fruit Trees, as they represent more
or less accurately the remote ancestral forms from which Citrus
arose. Every effort has been made to discover all possible
characters that serve to demonstrate the evolutionary progress and present
relationships of these plants. For this reason tens of thousands of
serial microtome sections were made by the modified Juel technique from
herbarium specimens. The new characters thus revealed have made
possible the taxonomic placement of many genera and species up to now only
imperfectly known, and sometimes known only from a single scanty herbarium
specimen. For example, in all the herbariums of the world, the new
genus Clymenia, closely related to Citrus, and the new species Citropsis tanakae were each represented by a single specimen,
each having only a single flower bud. Nevertheless, by application
of the modified Juel technique, it was possible to classify both of these
plants with certainly.
Thanks to this minute
and detailed study of all known plants of the subtribe Citrinae, a study that
has extended over many years, it became possible to discuss, with all pertinent
evidence assembled, the phylogeny of the True Citrus Fruit Trees, a problem of
great scientific interest because of the highly specialized pulp-vesicles
filled with fragrant, delicious juice. In the orange subfamily
these organs are found only in the subtribe Citrinae and in full perfection
only in the genus Citrus; nor are they known to occur in any other of
the half-million or so species of higher plants now recognized by taxonomic
botanists. Keys to the groups and genera of the subtribe Citrinae are presented.
GROUP A. THE PRIMITIVE CITRUS
FRUIT TREES
This group includes the five
genera of the subtribe Citrinae, Severinia, Pleiospermium,
Burkillanthus, Limnocitrus, and Hesperethusa, which show
the most primitive pulp-vesicles, lacking the stalks of the pulp-vesicles of
group C (the True Citrus Fruit Trees) and the definite conical shape with broad
sunken bases of those of group B (the Near-Citrus Fruit Trees).
Until three decades ago, taxonomists had not
realized that the plants belonging to these five genera contained in their
fruits various primitive forms of pulp-vesicles, those very remarkable
structures heretofore known only in the True Citrus Fruit Trees, where they
attain their most perfect development, and in the obviously related Near-Citrus
Fruit Trees, where they are somewhat simpler but still impossible to
overlook. As a result of this discovery, made during the original
preparation of this chapter, these five genera were transferred to the subtribe
Citrinae and took their place among the near relatives of
Citrus.
Severinia has six
species, all with simple leaves and with a cup-shaped disk in which the base of
the ovary is partially immersed. The very primitive pulp-vesicles are peripheral and much like those of
Hesperethusa described below.
Pleiospermium has
five species: one with trifoliolate leaves, rarely with two leaflets or even
one; another species with usually unifoliolate or difoliolate leaves, rarely
with three leaflets; the third, fourth and fifth species always with
unifoliolate leaves. All these species have winged petioles that
are rather long (one-eighth to one-sixth the length of the leaf blade) and
articulated with the blade. The locules of the fruit contain
numerous small, ovoid or cylindrical pulp-vesicles, with bluntly rounded or
acute tips, usually having an outer cortical shell of coarser tissue surrounding
a central core of more delicate tissue that seems to disintegrate, as the fruit
ripens, into a more or less spongy, oily, or resinous
mass.
Burkillanthus looks like a
Pleiospermium with giant, hard-shelled fruits. The uni-, di-,
or trifoliolate leaves resemble those of Pleiospermium dubium but are
larger; the petioles are narrowly winged and articulated with the leaflets, as
in all the species of Pleiospermium. The flower is larger
than in Pleiospermium and the ovary is very different in morphology,
having 22 to 26 ovules in each of the five-locules, the largest number known in
any plant of the orange subfamily. The fruits of
Burkillanthus are much larger than those of Pleiospermium and also
differ in having a firm leathery or woody rind. They contain very
numerous, very large seeds and peculiar thick-walled pulp-vesicles that seem to
secrete a mucilaginous fluid. Burkillanthus is probably
descended from a Pleiospermium-like ancestor but has become so different
as to constitute a very distinct genus.
Limnocitrus also resembles Pleiospermium slightly but differs in
having thick, veiny, simple leaves with very short petioles (one-tenth as long
as the leaf blade) that are pulvinoid for their whole length. The
ovary differs strikingly from that of Pleiospermium. The
fruits of Limnocitrus differ from those of Pleiospermium in having
very numerous, very slender, fusiform pulp-vesicles contracted at the base into
a short, broad stipe; they also show a central core of whitish droplets of oil
(or wax?). These pulp-vesicles do not show the clearly corticated
structure of those of Pleiospermium, or the irregular outlines of those
of Hesperethusa, which are also much larger and
fewer.
Hesperethusa presents striking
analogies in its leaf characters with Citropsis, the African
cherry-oranges. The pulp-vesicles in Citropsis are, however,
much more highly organized than those of Hesperethusa, and show a high
specialization of both structure and arrangement in the locule. The
pulp-vesicles of Hesperethusa
are irregularly rounded or even polygonal from natural pressure, without
specialization of form or regularity of arrangement as far as can be learned
from the material now available for study. Hesperethusa, like
Citropsis, can be grafted on Citrus, and vice versa.
This group of Primitive Citrus Fruit Trees
offers promise of throwing light on the origin and evolution of the
pulp-vesicles, unique organs that are found only in the subtribe Citrinae and
that reach their highest development in the genus Citrus and the other
closely related genera comprising the True Citrus Fruit Trees.
In the course of a comparative study of the
vascular traces of the flowers of all obtainable genera of the orange
subfamily, Tillson (1938, pp. 12 and 25; see also Tillson and Bamford, 1938,
pp. 787 and 791) found that all the genera comprising the Primitive Citrus Fruit
Trees show fusion of the lateral sepal bundles with the petal
midrib. This unusual type of floral vascular anatomy is found
elsewhere in Luvunga and Paramignya, in the species of
Citrus belonging to the subgenus Citrus (comprising all the
commonly cultivated citrus fruit trees), and in Swinglea.
XIV. Severinia
Tenore
XIV. Severinia
Tenore, Ind. Sem. Hort. Neap. 3. 1840; also in Atti Terza Riunione Degli Sci.
Ital. 502. 1841. Helie Roem. Syn. Hesper. 1:42.
1846.
Type species.—"Citrus
buxifolia Hort." = C. buxifolia Poir., in Lamarck, Encycl. Meth.
4(4):580 (1797) = Severinia buxifolia (Poir.) Tenore, loc.
cit.
Distribution.—Southern China,
Malay Peninsula, East Indian Archipelago to the Philippines, Kei Islands, and
New Guinea.
Common name.—Box-orange or
severinias.
Leaves simple, conspicuously
parallel-veined, with very short wingless petioles 1/15-1/10 the length of the
leaf blade, not articulated with it; flowers small; in simple clusters in the
leaf axils or in corymbs or panicles; sepals 3-5-lobed; petals 3-5; stamens
6-10, free; disk cup-shaped, enclosing the base of the ovary; ovaries with 1-5
locules, with 1 ovule pendent in each locule; fruits small, berry-like and
juicy or semidry with a well-defined peel dotted with oil glands; locules with
rudimentary pulp-vesicles, polyhedral through mutual pressure, often crushed by
the developing seeds; seeds smooth, plump, single in each locule.
The types [sic] species and all the other
species here referred to as Severinia have in recent decades been
considered to be species of Atalantia. Atalantia,
however, differs decidedly from Severinia in having well-formed, conical
pulp-vesicles and much larger flowers, often with the filaments of the stamens
more or less connate. The typical species of Severinia have
the ovary more or less sunken in the disk, which is, in consequence, usually
broader than the ovary. As noted above, Severinia has very primitive, stalkless, peripheral
pulp-vesicles very like those of Hesperethusa; in neither genera is there
indication of the contents breaking down into a more or less resinous mass, such
as is seen in Pleiospermium. Severinia differs decidedly from
Hesperethusa in having simple leaves and wingless petioles.
The leaves of Severinia are strongly veined, the finer veins forming more
or less uniform reticulations, not clearly marked in
Hesperethusa. A still more striking difference is in the
disk, which is cup-shaped in Severinia and annular in
Hesperethusa. Severinia resembles Atalantia in the
shape, size, and venation of its leaves and in having wingless petioles, but
differs decidedly in having nonarticulated petioles and more primitive
pulp-vesicles without definite form, those of Atalantia being conical and
pointing toward the center of the fruit with a rather broad base, more or less
sunken in the tissues of the dorsal locular walls. Severinia
also resembles somewhat several of the genera in the subtribe Triphasiinae,
especially the genera Paramignya and Pamburus, from which it
differs in possessing rudimentary
pulp-vesicles.
Severinia lauterbachii
is very different from the first five species of the genus and is as yet
only imperfectly known.
Severinia buxifolia
and S. disticha are known to have served as rootstocks for
Citrus, and it is probable that other species can be so employed.
The six species of Severinia are keyed.
1.
Severinia buxifolia (Poir.) Tenore, Ind. Sem. Hort. Neap. 3 (?). 1840;
also in Atti Terza Riunione Degli Sci. Ital. 501-503. 1841. Citrus
buxifolia Poir. in Lamarck, Encycl. Meth. 4:580. 1797; Limonia
monophylla Lour. (non L.) Fl. Cochinch. 1:271. 1790; Citrus
emarginata Desf. 1829; Limonia bilocularis Roxb. 1832;
Sclerostylis atalantioides Wt. & Arn. Prodr. 93. 1834; Atalantia
loureiriana Roem. Syn. Hesper. 1:44. 1846;8 Helie atalantioides
(Wt. & Arn.) Roem. Syn. Hesper. 1:42. 1846; Sclerostylis
buxifolia Benth. Hook. Jour. Bot. 3:326. 1851; Atalantia
buxifolia Oliv. Jour. Linn. Soc. 5 (Suppl. 2):26. 1861; Severinia
monophylla ("L.") Tan. Bul. Mus. Hist. Nat. Paris, 2 ser. 2:163.
1930.9 Illus. Seemann, Bot.
Voy. Herald, pl. 81. 1852-1857; Penzig, Studi Bot. Sugli Agrumi, Atl. pl. 11,
figs. 6-17; pl. 12, figs. 1-21. 1887; Swingle, Jour. Wash. Acad. Sci.
5:656, 657, figs. 1, 2. 1916; Tillson & Bamford, Amer. Jour. Bot.
25:784, figs. 21-25. 1938; fig. 3-18 this work.
Type
locality.—In the vicinity of Canton, China (Sonnerat). Herb.
Mus. Nat., Paris.
Distribution.—Southern China: Kwangtung Province, Hainan Island; North
Vietnam; South Vietnam; Taiwan; Hong Kong; Cambodia;
Laos.
Common name.—Chinese
box-orange.
Bentham's description (1851, p.
326) may be translated as follows: "Low shrub, or in our gardens dwarf tree,
branches spreading, young twigs obscurely angular or somewhat compressed,
always puberulous, soon glabrous and terete. Spines axillary,
strong, usually much shorter than the leaves, only rarely longer.
Leaves 1-1 1/2 in. [2.5-3.8 cm] long, 6-9 lin. [12-18 mm] wide, obtuse,
narrowed at the base into a short, narrow, rounded petiole, coriaceous, rigid,
densely dotted with oil glands, glabrous, with numerous parallel veins arising
from the midrib, much more conspicuous than in many species [of Sclerostylis
= Atalantia]. Flowers between small bracts in the axils
of the leaves, sessile, or rarely borne on a very short pedicel, never
solitary, usually 2-3-fasciculate. Calyx widely campanulate, short,
smooth, with 5 rounded, very obtuse lobes. Petals 5, scarcely 2
lin. [4 mm] long, oblong, erect, glabrous. Stamens 10, shorter than
the corolla, filaments dilated, attenuate at the apex, anthers ovate, parallel
locules. Ovary sessile on a cupulate disk, subglobose, fleshy, with
small locules in the centers, ovules attached to the central axis slightly
below the apex. Style very short, stigma thick, ovoid,
pulvinate. Berry depressed globose, becoming black at maturity,
seeds large, ovoid."
The original description
of Poiret (1797, pp. 580, 581) reads, in translation, as follows: "Citrus
buxifolia, leaves subsessile, ovate-retuse, flowers racemose, very small…I
do not know its height, but the branches I have examined in the herbarium of
Citizen Lamarck, would indicate that it is not tall. With its
short, stiff branches, its spines and the shape and firmness of its leaves, it
has the aspect of Rhamus pyracantha. The wood is hard,
white, with a smooth green bark. It has many spreading branches
with stiff straight spines, yellowish at the tips. The leaves are
few, alternate, oval, almost sessile, much like those of box, but twice as
large, obtuse at the apex, emarginate, narrowed at the base, remarkable by the
very close veins, which are prominent and parallel, coriaceous, with entire
margins. The petioles are simple and very short. The
flowers occur in small bunches toward the ends of the branches. The
corolla is white and very small. This plant is native to China and
was observed there by Sonnerat, who sent the specimens to Citizen Lamarck."
This, the type species of the genus
Severinia, is not an uncommon plant in wooded areas in southern China as
far north as the Tropic of Cancer. It is also found in Hong Kong and
Hainan, and in Indo-China from the northern border of Laos and North Vietnam
to latitude 10° S. It is a small, spiny tree with crooked, thorny
branches and dense, evergreen foliage (see fig. 3-18). It is esteemed by the Chinese for its
leaves, which are used in making yeast cakes. For this reason it is
called 'tsau 'ping lak10 in Cantonese, meaning "wine
cake thorn."
A careful search for variations
in S. buxifolia, made by G. W. Groff and F. A. McClure, formerly of the
Horticultural Department of Lingnan University at Canton, led to the discovery
of many forms of the species differing in height, character of growth, size and
shape of the leaves, number and length of spines, et cetera, and also in
physiological characters, such as congeniality with Citrus in grafting
experiments. So far, none of these forms have been given taxonomic
names, but some appear so distinct that when they are studied more carefully it
may prove desirable to recognize them as subspecies, varieties, or forms.
Certain broad-leaved forms growing near the
seashore at Da Nang (Tourane) in South Vietnam (Lat. about 16° N.) have shown
not only great vigor under culture but also high resistance to boron and
injurious saline solutes often found in the ground water where Citrus is
grown in semiarid situations.
Inasmuch as
Citrus can be grafted readily on S. buxifolia, and since some
plants so grafted have lived over thirty years without showing any sign of bad
graft union, it is clear that more study should be given to the use of this
species as a rootstock. Besides its important physiological
distinction from Citrus in respect to boron absorption and sensitivity toward
saline solutes in the ground water, the very fact that S. buxifolia is
so remote a relative of Citrus as to be quite immune to many Citrus
diseases recommends it for trial as a rootstock.
With regard to the relative boron tolerance of
the lemon and the box-orange, Eaton and Blair (1935, p. 413) stated: "The lemon
(Citrus limonia Osbeck) is highly sensitive to boron; in fact few plants
are thought to be more sensitive, and it accumulates much boron in its
leaves. The Chinese box orange, Severinia buxifolia, on the
other hand, in confirmation of the observations of Swingle, Robinson, and May,
is relatively tolerant to boron. Severinia has been found to
accumulate much less boron in its foliage than does the lemon."
Eaton and Blair conducted experiments on boron
tolerance of the lemon and of the Chinese box-orange, as well as of
reciprocally grafted plants (i.e., lemon on box-orange roots and box-orange on
lemon roots) which Swingle supplied them with the help of Eugene May,
Jr. Striking proof that the box-orange has much higher resistance
to boron-poisoning than the lemon is given in their Table II reproduced below
(table 3-2).
Eaton and Blair
commented as follows on the data presented in the table: "Lemons [grafted] on
box orange roots had about one-third as much boron in their leaves as when
grown on their own roots. The data indicate that the boron
concentrations in the box orange leaves are increased threefold when the plant
is grown on lemon roots. There were no box orange plants on their
own roots in the 2 and 4 ppm cans, but the plant in 6 ppm on its own roots had
390 ppm, whereas the box orange on lemon roots in 4 ppm had 877 ppm of boron in
its leaves. Lemon leaves on their own roots were more severely
injured than when on the box orange roots."
This record of experiments, made after years of experience in growing
Citrus in nutrient solutions containing known amounts of boron, is of
great value in showing the striking difference in the toxicity of boron for
Citrus roots and for Severinia roots, although both genera of
plants grow wild in situations that are similar with respect to both climate
and soils. (See also discussion of boron tolerance of Eremocitrus.)
The possibility of growing Citrus on a
plant as remote, taxonomically, as the box-orange has served to direct
attention to the need of testing all the wild relatives of Citrus in the
hope of finding others able to thrive as the box-orange does, in physical and
biological environments that Citrus roots cannot endure, and at the same
time able to serve as rootstocks for the commonly cultivated species of
Citrus.
1a. Severinia buxifolia brachytic
form.
Common name.—Dense-leaved
box-orange.
Twigs spineless or nearly so,
internodes very short, often less than 1 cm long; leaves densely crowded
because of the short internodes; flowers and fruits as in the species but
produced in less abundance.
This form,
analogous to the myrtle-leaved variety of the sour or Seville orange, Citrus
aurantium var. myrtifolia, is even more like the common box (Buxus
sempervirens) in general appearance than the normal form of the Chinese
box-orange. This dense-leaved variety is, like the parent species,
able to endure high temperatures, hot winds, and other unfavorable climatic
conditions that would injure or kill the true box tree. It cannot,
of course, endure severe cold, like the common box, and is injured when the
temperature falls as low as 14° F (-10° C). This beautiful, vivid
green, nearly thornless shrub deserves careful trial as a substitute for the
common box for hedges in warm-temperature and subtropical climates.
This form seems to arise by mutation from the
mother species. As might be expected, intermediate forms occur as,
for example, the one shown in figure 3-18, which has some nearly thornless branches and others
with long spines borne on the same plant. Even the typical forms of
this mutation are usually not entirely thornless but have short, slender
spines that are, however, so completely hidden under the very abundant leaves
that the spines are scarcely felt when a branch is touched.
Doubtless by careful selection superior, dense-leaved, thornless forms of this
variety can be developed that will be of high ornamental value.
2. Severinia disticha (Blanco)
Swing. Jour. Wash. Acad. Sci. 28:533. 1938. Limonia disticha
Blanco, Fl. Filip. 356. 1837; L. corymbosa Bl. Fl. Filip. ed. 2. 251.
1845; Sclerostylis nitida Turcz. Bul. Soc. Nat. Mosc. 31:249.
1858; Atalantia nitida Oliv. Jour. Linn. Soc. 5 (Suppl. 2):25.
1861; A. disticha (Blanco) Merr. Phil. Gov. Lab. Bur. Bul. 27:28.
1905.
Type.—Wanting.
Substitute type: Luzon (Merrill, Species Blancoanae, No.
594). Herb. Bur. Science,
Manila.
Distribution.—Philippines:
Luzon to Mindanao islands; Sabah (North Borneo): Banguey
Island.
Common name.—Philippine
box-orange.
Young twigs angular, sometimes
with scanty gray pubescence soon glabrous and terete, sometimes apparently
glabrous from the beginning; twigs spineless but occasionally with paired,
slender, sometimes pubescent, spine-like paraphylls rarely equaling the
petioles in length; leaves 3-8 (rarely 10-11) cm long by 2.5-4 (rarely 1.5-2)
cm wide, oval, ovate-lanceolate or sometimes fusiform, acuminate or acute at
the apex, acumen often blunt and frequently emarginate, narrowed gradually into
a cuneate base which merges into the petiole, margin subentire sometimes
slightly crenulate and frequently wavy in dried specimens, lateral veins
visible above but slightly more prominent below, extremely numerous, parallel,
straight, occasionally showing 15-20 pairs, arising at an angle of 55°-60° with
the midrib; petioles 3-9 (usually 6-8) mm long, flattened above with a furrow
made by the decurrent margins of the leaf, sometimes pubescent, sometimes
almost glabrous, especially on the under side; inflorescences many-flowered,
axillary or rarely terminal racemes, 3-6 cm long (rarely 8-9 cm long, and much
branched); pedicels slender, 3-5 mm long, with minute bractlets at their
juncture with the peduncle; both peduncles and pedicels pubescent; calyx
persistent, sepals 5, imbricate, broadly rounded, margins ciliate; petals 5,
3.5-4.5 mm long, 2-3 mm wide, oblong or obovate; stamens 10, nearly as long as
the petals, filament flattened, broad, free, anthers short, small, ovate; ovary
subglobose, small (1.5-2 mm long), 2-locular, with 2 large glands at apex, 1
over each locule; disk cupulate, partly enclosing the base of the ovary; style
as long or longer than the ovary, stigma ovoid, slightly broader than the
style; fruits subglobose, 6-8 mm diam. when mature, peel yellowish-green, with
many large, translucent oil glands, 0.5-0.75 mm diam.; seeds thick,
monoembryonic, testa exceedingly thin, embryo olive-green when dry and covered
with numerous minute oil glands, 0.1 mm diam.
This species, very abundant in the northern Philippine Islands, is so like the
box-orange, S. buxifolia, that when it was found the latter could be used
successfully as a rootstock for Citrus tests were also made with S.
disticha. It was found to serve as a rootstock almost as well as
the box-orange and, like it, to be resistant to amounts of boron in the soil
moisture that would be injurious to Citrus grafted on the commonly used
rootstocks belonging to the genus Citrus. Swingle made these
tests over thirty years ago in a greenhouse at Washington, D.C. by watering
potted seedling plants of Citrus, Severinia, and other genera of
the orange subfamily with water containing 2 ppm of boron. Because
of evaporation from the soil and transpiration from the plant, the boron
accumulated in the soil rather rapidly and soon the Citrus seedlings
showed the characteristic yellowish-green mottling of the leaves, followed by
severe injury and even death. The seedlings of Severinia
buxifolia and S. disticha remained healthy for many weeks after those
of the Citrus species showed severe injury. Severinia
disticha has fruited freely in Miami, Florida.
3. Severinia linearis (Blanco)
Swing. Jour. Wash. Acad. Sci. 28:533. 1938. Limonia linearis
Blanco, Fl. Filip. 357. 1837; Atalantia linearis Merr. Phil. Jour. Sci.
1 (Suppl. 3):200. 1906.
Type.—Wanting. Substitute type: Luzon, Rizal Prov.
(Merrill, Species Blancoanae, No. 746). Herb. Bur. Sci., Manilla
[sic].
Distribution.—Philippines: Luzon
Island.
Common name.—Narrow-leaf
box-orange.
Merrill's description of
Atalantia linearis reads as follows: "A shrub 1 to 3 m high.
Branches light gray, glabrous, the young branchlets greenish, pubescent or
puberulent, terete. Leaves alternate, linear to narrowly
linear-oblong, or linear-lanceolate, glabrous, shining, coriaceous, 2 to 7 cm
long, 3 to 10 mm wide, the base acute, the apex blunt, retuse, entire, the
margins often somewhat recurved, the midrib prominent, the lateral nerves
numerous, scarcely more distinct than the dense reticulations; petioles
glabrous or pubescent, 3 mm long or less. Inflorescence of terminal
and axillary panicles, 3 to 5 cm long, pubescent, the branches short,
few-flowered. Flowers white, short-pediceled, and 6.5 mm
long. Calyx short, regularly 5-lobed, the lobes imbricate, broadly
suborbicular, rounded, about 2 mm long, 2.5 mm wide, pubescent, the margins
ciliate. Petals 5, free, oblong, the apex rounded, 6 to 6.5 mm long,
and 3 mm wide, somewhat narrowed at the base. Stamens 5 [10];
filaments broad, 4 mm long; anthers about 1-4 mm long. Ovary
glabrous, 3-celled, sessile, the disk shallow. Fruit glabrous,
globose, white, glandular-punctate, about 8 mm in
diameter.…
"On cliffs and boulders at an
altitude of about 40 m along the river, frequently in situations submerged at
high water associated especially with Eugenia mimica Merr., and
sometimes with Homonoya riparia Lour.…A species at once recognized by its
very narrow leaves, which are retuse at the apex. Blanco's original
description of this species is very short, his material being from the Island
of Maricaban, Province of Batangas, Luzon, flowering in the month of
July. In this description he states that the leaves are minutely
serrate, probably from the fact that in specimens with recurved leaf margins,
the slightly raised veins on the upper surface appear like minute
teeth. The note following the description of the species in the
first edition of the Flora di Filipinas is excluded in the second, this
note referring to a similar form observed by Blanco in the Province of Bulacan,
Luzon. I am of the opinion that the form here described is
identical with Blanco's Limonia linearis, and accordingly his specific
name is adopted and the species is redescribed under Atalantia."
This curious species is apparently a satellite
species derived from S. disticha that has become adapted to growing in
situations exposed to flooding. Perhaps the narrow leaves resist
the tearing action of flood waters rushing down narrow valleys. The
lateral veins of the leaves run less regularly parallel than those of the
leaves of S. disticha. Perhaps this, too, is an adaptation to
prevent splitting of the leaves by rushing waters.
Extreme specimens of S. linearis are
easily distinguished by the narrow leaves, but broad-leaved forms are hard to
distinguish from narrow-leaved forms of S. disticha. A
broad-leaved specimen of S. linearis (coll. by M. P. Sulit, April, 1926;
Tanaka’s ident. No. C-305, Forestry Bur., No. 30320, Herb. Univ. Calif.) has
leaf blades measuring 9 to 11 cm in length, and 1.5 to 2 cm in
width! However, the venation of the two species differs slightly,
lateral veins of S. linearis running more irregularly and distinctly less
parallel than those of S. disticha. It is possible that
natural hybrids may be in play here.
4. Severinia paniculata (Warb.) Swing. Jour. Wash. Acad. Sci.
28:533. 1938. Atalantia paniculata Warb. Bot. Jahrb.
13:340. 1891; A. maritima Merr. Phil. Jour. Sci. Bot.
9:293. 1914; A. disticha var. paniculata Tan. Jour. Arnold
Arbor. 9:141. 1928.
Type.—Ceram
Laut (Warburg, No. 20132). Herb. Bot. Mus.,
Berlin-Dahlem.
Distribution.—Moluccas:
Kei Islands, Ceram Laut; southern Philippines: Mindanao, Negros, and Panay
islands; North Borneo (Sabah): Biak; Sumbawa and Java (fide Tanaka,
1928c, p. 141).
Common
name.—Bouquet box-orange.
Warburg's
original description of this species reads in translation as follows: "A shrub
with gray-pubescent [terminal] branches, soon glabrate, with yellow wood;
leaves oblong-ovate, obtuse or emarginate at the apex, margin obscurely
crenulate; petioles short, pubescent, nearly terete; inflorescences terminal,
paniculate, pubescent, pedicels short; calyx 5-lobed, lobes pubescent and
ciliate, subtruncate at the tips; petals nearly glabrous, lanceolate, obtuse;
filaments long, glabrous, anthers broadly cordate; ovary with 4 locules; style
deciduous; fruits globose, with the calyx persistent, 2-4-seeded.
The youngest twigs are reddish-yellow, the somewhat older twigs brown after the
gray pubescence has fallen, often with a whitish bloom; the petioles are 8 mm
long, the leaves 9-12 cm long and 3-5 cm wide, with a large number of parallel
veins of which some are more prominent, uniting near the margins in
arches. The whole inflorescence is 4-6 cm long, branched from the
base, the flower pedicels are from 2-4 mm long, the calyx is about 2 mm long,
the sepal tips 1 mm long, the petals and stamens are 4 mm long; the largest
fruit of my specimen [Warburg, No. 20132] is 12 mm in diameter."
This species is easy to distinguish from the
related species because of its pubescence, its lack of thorns, and its terminal
paniculate inflorescences.
Severinia
paniculata was discovered on Ceram Laut and Kei Islands in the Moluccas
west of New Guinea growing on dry areas in the brush. Tanaka
considered it to be identical with Atalantia maritima, growing in the
southern Philippines, but Swingle could not follow him in making the species a
variety of S. disticha, although it is closely related to that
species. It differs in having larger leaves and smaller, paniculate
rather than racemose inflorescences, and in having ovaries with four instead of
two locules.
5. Severinia
retusa (Merr.) Swing. Jour. Wash. Acad. Sci. 28:533. 1938.
Atalantia retusa Merr. Phil. Jour. Sci. 1(Suppl. 4):200.
1906.
Type.—Philippines, Palawan
Island, Puerta Princessa (Curran, No. 3609). Herb. Bur. Sci.,
Manila.
Distribution.—Known only from
the type locality.
Merrill's original
description of this species reads as follows: "A shrub about 3 m high, with
oblong-elliptical-ovate, subcoriaceous, glabrous, strongly retuse leaves,
racemose inflorescences and 5- to 7-merous, the stamens 10 to 15, free, the
ovary 1-celled. Branches brownish-gray, glabrous, the branchlets
green, glabrous. Leaves 5 to 9 cm long, 2.5 to 4 cm wide, shining,
broad at both ends, scarcely narrowed above, the base rounded; nerves numerous,
anastomosing; petioles 8 mm long or less, the spines short or
wanting. Racemes in the upper axils, 1.5 cm long in anthesis,
densely flowered, puberulent. Calyx lobed, the teeth 5 to 7, short,
broad, regular, their margins ciliate, obtuse. Petals 5 to 7,
glabrous, oblong, obtuse, 5 to 5.5 mm long, 2.5 mm wide. Stamens
10 to 15, unequal, the filaments free, 2.5 to 3.5 mm long, anthers broadly
ovate, slightly exceeding 1 mm in length. Ovary oblong, glabrous,
1-celled, 1-ovuled; style much shorter than the ovary, including the stigma
about 1 mm long. Disk thickened, ring formed. Fruit
(immature) ovoid, glabrous.
"A species
apparently related to Atalantia disticha (Blanco) Merr., differing from
the latter in its leaves not being narrowed above, and in its flower
characters. In some cases a very rudimentary second cell was
observed in the ovary, showing that the species is perhaps sometimes with
2-celled ovaries. The very short style is another character, as well
as the variable number of calyx teeth, petals and stamens. In other
species of the genus the ovary is from 2- to 5-celled, the petals and calyx
teeth 3 to 5, and the style equalling [sic] or longer than the
ovary."
This species is described as having an
ovary with only one locule, but a specimen in the National Herbarium at
Washington, sheet No. 439318, from Palawan (J. Bermejos, Bur. Sci., No. 195),
shows ovaries with two locules. Penzig (1887, p. 158) reported
finding, very rarely, ovaries of S. buxifolia that were "monocarpellate
without the slightest trace of a second ovarial follicle." A
specimen of Atalantia maritima (considered by Tanaka [1932e, p.
426] to be a synonym of S. paniculata) in the National Herbarium at
Washington, sheet No. 1294389, from Zamboanga, Mindanao Island, P.I. (A.
Lomente, Forestry Bur., No. 25288), shows only a single carpel in the ovary and
a single stylar canal. Three of the typical species of
Severinia occasionally (or regularly) show only a single locule in the
ovary and probably the other closely related species, S. paniculata, may
occasionally show a hypomerous or even a monocarpellate gynoecium.11
SPECIES OF UNCERTAIN RELATIONSHIPS
6. Severinia lauterbachii Swing. Jour. Wash. Acad. Sci.
28:533. 1938. Atalantia litoralis Lauterb. (non Guill.) Nova
Guinea 14:146. 1924.
Type.—Northwestern New Guinea [West Irian, Indonesia], Schouten Islands,
Biak, near Warsa (Lat. 2° S., Long. 137° E.), growing on coral limestone strand
(Feuilleteau de Bruyn, No. 320). Herb. Bot. Mus.,
Berlin-Dahlem.
Distribution.—Known only
from the type locality.
Common
name.—Triangle-fruit severinia.
The
original description, translated, reads as follows: "Small tree, about 4 m
high, with slender, flattened, glabrous twigs, 4 mm thick. Leaves
elliptic or broadly lanceolate, subacutely rounded at the apex, sometimes
emarginate or subacute, base subacute, margins obscurely crenulate, 12-16 cm
long, 6-8 cm wide, coriaceous, glabrous above and below, lateral veins 16,
obliquely subparallel, curved and anastomosing near the margin, veinlets
subparallel, reticulate, when dry more prominent below like the midrib; petiole
flattened on upper surface, 6-7 mm long. Inflorescences axillary,
paniculate, many-flowered, 3-6 cm long, small branchlets minutely
pubescent. Flowers pedicellate, pedicels 3-4 mm long; buds obovoid;
calyx 5-lobed, lobes rounded, ciliate, 1 mm long; petals 5, oblong, rounded,
white, glandular, glabrous, margins subpellucid, 5 mm long, 1.5 mm wide;
stamens 10, 5 of them shorter, 3 or 5 mm long, both [lengths] connate at the
base, filaments dilate, glabrous anthers cordate, about 1 mm long; disk short,
10-crenulate; ovary triangular, each angle crowned with a large gland,
3-locular, 1.3 mm long; style clavate, 2 mm long, stigma decurrent, somewhat
3-lobed."
This species was said by Lauterbach
to differ from Atalantia paniculata Warb. (here S. paniculata) in
having much larger leaves and larger flowers with triangular ovaries, each
having three locules. The term "10-crenulate" applied to the disk
of this species probably indicates that it is cupulate (as in other species of
Severinia), but in drying it has become separated from the base of the
ovary and acquired a wavy margin. It is possible that this curious
species does not belong to the genus Severinia, but it is impossible to
place it with certainty until the flowers and fruits are better
known.
Merope angulata, widely
distributed in tidal swamps, also has triangular (sometimes 4- or 5-angled) fruits,
but it has very large seeds, stout paired spines in the leaf axils, an ovary
with two to four pendent ovules in each locule, and a cylindric disk definitely
not cupulate or 10-crenulate. Merope angulata is probably
very different from S. lauterbachii, although the latter species is also
a strand plant with triangular fruits.
This
species was named Atalantia litoralis by Lauterbach in 1924, but this
name was preoccupied by Atalantia littoralis (Miq.) Guill., a new
combination made eleven years previously by Guillaumin (1913, p. 441) when he
transferred Paramignya littoralis Miquel to the genus
Atalantia. The variation in spelling, litoralis or
littoralis, does not make these names distinct, as these are merely
variant spellings of the same Latin adjective. Lauterbach's
specific name, therefore, has no standing and must be supplanted by the new name
which is used here.
XV. Pleiospermium
(Engl.) Swing.
XV.
Pleiospermium (Engl.) Swing. Jour. Wash. Acad. Sci. 6:426. 1916.
Limonia Sec. Pleiospermium Engler, Die Nat. Pflanzenfam.
3(4):189. 1896.
Type
species.—Limonia alata Wt. & Arn. = Pleiospermium alatum
(Wt. & Am.) Swing.
Distribution.—Southern India, Ceylon, Sumatra, Java, Borneo.
Common name.—Orangeasters.
Small trees; twigs with 1 or 2 spines in the
axils of the leaves or unarmed; leaves 3-foliolate, 2-foliolate, or
1-foliolate, margins entire; petioles narrowly winged or nearly wingless
(broadly winged in P. latialatum); inflorescences axillary or terminal
panicles of few-flowered groups of flowers arising in the axils of the leaves;
flowers 12-24 mm diam.; calyx 4-5-lobed, sepals deltoid or lanceolate-acuminate
(in P. longisepalum, long, strap-shaped, persistent, reflexed); petals 4
or 5, oblong or linear-oblong, rounded at the tips, 6-12 mm long; stamens 8 or
10 (twice as many as the petals), filaments free, glabrous; anthers oblong or
linear-oblong; pistil subsessile or short-stalked; disk cupulate, enclosing
base of ovary for 0.5-0.8 mm; ovary cylindric or ovate, with 4-5 locules,
ovules 2 in each locule; style slender or thick, not sharply delimited from the
ovary, ending in a more or less capitate stigma with 4 or 5 stylar canals, with
2 small oil glands between each pair; fruit globose or oblong, 20-30 mm diam.,
with slender rudimentary pulp-vesicles, 2-10 mm long, that are more or less
corticated and show more or less disintegration of the contents into an oily or
resinous mass as the fruits ripen; seeds about 1 cm long, flattened.
This genus includes some of the most puzzling
species of the subtribe Citrinae. These species have been referred
to several different genera, usually with a question mark! None
have been examined thoroughly in the living condition and it is very desirable
that a study be made of the structure of the fruit and in particular of the
peculiar pulp-vesicles, which at some stages of their development have very
much the appearance of stalked oil glands.
The two always unifoliolate species from northeastern Borneo, P.
longisepalum and P. latialatum, and the one from Sumatra, P.
sumatranum, are somewhat like Atalantia in appearance, but differ in
having well-developed winged petioles plainly articulated with the leaf blade,
as in the two other species of Pleiospermium which have tri-, di-, or
unifoliolate leaves. The very curious new genus Burkillanthus
has a marked superficial resemblance to Pleiospermium in its leaf
characters but differs widely in its fruits. It is probably a
large-fruited form derived from a Pleiospermium-like
ancestor.
The pulp-vesicles in the genus
Pleiospermium, although very primitive in comparison with those of
Citrus, show a very interesting series of stages in the evolution of
these unique and taxonomically highly important organs. In P. dubium the pulp-vesicles are long-ovoid or cylindrical,
each with a very distinct tissue surrounding a definitely limited central body
that looks very much like a rutaceous oil gland, and, like such an oil gland,
degenerates at length into a granular, apparently structureless mass of oily
or resinous (?) substance. In P. sumatranum the
pulp-vesicles are short, only about 1 to 2 mm long, with blunt tips, and also
are plainly corticated. The central mass of cells breaks down into
an unorganized mass that looks much like that of an oil gland in the immature
peel of a citrus fruit. The pulp-vesicles of this species seem to
secrete a liquid that fills the locules of the fruit and hardens on drying to a
dark reddish-brown, brittle, resinous mass. In the two Bornean
species, P. longisepalum and P. latialatum, the pulp-vesicles are
longer than in P. dubium and much longer than in the similar-looking
P. sumatranum, and are of a slender conical shape often ending in a sharp
point at the apex. Their cortical tissue is much thinner than in
P. dubium and less clearly cellular. The constituents do not
break down so completely. The pulp-vesicles of these two species
are more like those of Citropsis and Atalantia but have smaller
bases not so deeply sunken into the dorsal wall of the locule. The
series of pulp-vesicles in the different species of Pleiospermium connect
with the still more highly developed pulp-vesicles of Limnocitrus, which
are no longer conical but are definitely fusiform and do have a very short
stalk.
Perhaps no other genus of the orange
subfamily is of greater interest than Pleiospermium in throwing light on
the origin and development of the unique and highly specialized pulp-vesicle
organs which are found only in the subtribe Citrinae, and which reach their
highest specialization in the genus Citrus, giving the delicious flavor
characteristic of its cultivated varieties. It would be desirable
to study the development of the pulp-vesicles in Pleiospermium and the
related genera Limnocitrus and Burkillanthus from living
material.
The discovery of three species of
Pleiospermium having unifoliolate leaves greatly resembling those of
Citrus in shape, size, color, and texture brings them and their
trifoliolate congeners into prominent notice as possible persisting
representatives of ancient ancestral forms in the descent of
Citrus. Like the living species of Citropsis in
tropical Africa, the species of Pleiospermium show all stages in the
origin of unifoliolate leaves from trifoliolate leaves (or pinnate leaves in
Citropsis), something that without doubt occurred, ages ago, in the
development of Citrus. A key to the species of Pleiospermium is presented below.
1. Pleiospermium alatum
(Wt. & Arn.) Swing. Jour. Wash. Acad. Sci. 6:428. 1916. Limonia
alata Wt. & Arn. Prodr. 1:92. 1834. Illus. Wight, Ill.
Ind. Bot. 1:pl. 41. 1840; Beddome, Fl. Sylv. pl. 7, fig. 3(1-9).
1871.
Type.—Southwestern British India,
"foot of the Neelgherries," Madras State (Wight, No.
324).
Distribution.—Western peninsular
India: Madras State; Ceylon.
Common
name.—Ceylon orangeaster.
Small trees with
glabrous branchlets, sometimes with a single stout, straight spine up to 25 mm
long at the side of the bud in the axil of the leaf, sometimes spineless,
especially the fruiting branches; leaves 3-foliate [sic] (rarely 2- or
1-foliolate); lateral leaflets much smaller, all leaflets rather thick, glossy
above, finely netted-veined beneath, lateral veins slender, not very
conspicuous, making a very wide angle with the midrib, margins entire, the apex
obtuse or subacute, rarely acuminate, bluntly rounded at the very tip, often
emarginate, the leaflet blade abruptly narrowed to a cuneate base; petiole
variable, 25-38 mm long, narrowly winged, articulated with the blade;
inflorescences in clusters in the axils of the leaves or in terminal
much-branched hoary panicles; flowers small, about 12-15 mm diam., 4-5-merous,
pedicels rather short, finely pubescent; flower buds cylindric, rounded at the
tip, more or less pubescent; calyx small, 4-5-lobed, sepals deltoid, finely
pubescent; petals oblong or ovate, obtuse, entire, sparingly covered with fine
pubescence without; stamens free, 8 or 10 (twice as many as the petals),
filaments free, anthers large, erect, linear-oblong; pistil subsessile, seated
on a low cupulate disk which clasps the base of the ovary for 0.5-0.8 mm; ovary
obovoid, glabrous, 3.5-4.2 X 1.6-1.8 mm, 4-5 locular, with 2 ovules in each
locule; style slender, gradually merging with the tip of the ovary and capped
by a slightly thicker, subglobose stigma; fruits globose, 2.3-2.5 cm diam.,
like small oranges in appearance, the segments each containing 1 or 2 seeds
surrounded with an aromatic, mucilaginous fluid, peel rough, dotted with
numerous oil glands, seeds ovate, somewhat flattened, with a hard, smooth
testa.
This species occurs rather commonly in
the hot dry parts of southern India and Ceylon. The fruit is said
to be very bitter; the wood is hard and heavy, yellowish,
even-grained. The leaves have a strong lemon odor when
crushed.
2. Pleiospermium
dubium (Bl.) Swing. Jour. Wash. Acad. Sci. 6:429. 1916.
Limonia ? dubia Blume, Bijdr. Fl. Nederl. Indie 1:133.
1825; Paramignya blumei Hassk. Tijdschr. Nat. Gesch. 10:137.
1843.
Type.—Java, Kuripan (Blume),
Rijks Herb., Leiden (sheet No.
908203-1318).
Distribution.—Java.
Common
name.—Java orangeaster.
Twigs angular when
young, with straight, sharp spines, 2-2.5 mm long, 1 or 2 in the axils of each
leaf, older twigs often spineless; leaves 1-, 2-, or 3-foliolate; leaflets
thin, elliptical, cuneate at the base, acute or acuminate at the apex, the
very apex often blunt, or emarginate, the middle leaflet (or single leaflet)
7-9.5 X 2-3.6 cm, with 8-12 faint veins visible on both surfaces on each side;
petioles 7-22 mm long, narrowly winged above but nearly wingless at base,
plainly articulated at both ends; flowers in short, 5-15-flowered, axillary
racemes; ovary obovoid, hirsute, 2-2.8 X 1.2-1.7 mm, borne on a cylindrical
gynophore somewhat like that of Paramignya or Merope; fruits with
rudimentary pulp-vesicles arising from the ovary wall, elongate-ovoid or
irregularly conical, 2.5-3.5 mm long, 0.4-0.6 mm broad at the base, completely
covered with a cellular cortex which connects at the base with the dorsal wall
of the locule but is not usually imbedded in it and never deeply imbedded;
mature fruits subglobose, 2-2.5 cm diam.
This
species, known only from Java, is of great interest from the fact that, because
of its leaves, which have from one to three leaflets, it serves to connect the
usually trifoliolate Indian species, P. alatum, with the unifoliolate
species, P. samatranum, P. longisepalum, and P. latialatum,
which all have leaves remarkably like those of Citrus. This
is true also of the unifoliolate leaves of P. dubium.
The pulp-vesicles of this species are each
sharply divided into a cortical layer, composed of rather small cells, and a
central portion made up of larger cells which finally disintegrate into a
spongy mass, oily or resinous (?) in nature, which looks much like the contents
of the oil glands in the peel of the young fruit (see above).
Blume's type material
was collected on "limestone hills near Kuripan, Buitenzorg Prov." in Java, but
neither flowers nor fruits were mentioned by him. The type specimen
in the Rijks Herbarium at Leiden, No. 908203-1318, and a cotype specimen in the
Herbarium of the Museum of Natural History at Paris both show trifoliolate
leaves; the Leiden branch has paired axillary spines as described by Blume, the
Paris specimen is spineless.
Plants of P.
dubium cultivated in the Botanic Garden at Buitenzorg (now Bogor), Java,
have flowered and fruited for many years. Dried herbarium specimens
from these trees were distributed by B. P. G. Hochreutiner, as No. 111 in his
Plantae bogorienses exsiccatae, about 1904 (this number is found in the
Kew Herbarium and also in the Paris herbarium). On his printed
label Hochreutiner stated: "This plant is without any doubt the original of
Hasskarl['s Paramignya blumei]" (Swingle, 1916b, p.
429). Herbarium specimens from these Buitenzorg plants are
preserved in the herbarium of the Bureau of Science at Manila and in the
National Herbarium (sheet No. 448333) at Washington. From them were
taken the flower and fruit characters given in the description
above.
Pleiospermium dubium is a small
tree or shrub, attaining a height of about 4 meters. The
inflorescences were said by Koorders (1912, p. 426) to be 5 to 15-flowered,
short, axillary racemes. This plant has probably the most
polymorphic foliage of any species of the orange subfamily. It
often has branches bearing only trifoliolate leaves and others with exclusively
unifoliolate leaves. However, sometimes trifoliolate and
unifoliolate leaves are disposed hit-and-miss on a branch. Some
trifoliolate leaves develop only one of the side leaflets.
This species and Citropsis gabunensis of tropical West Africa, which has almost
equally polymorphic leaves, offer excellent material for a study of the
critical evolutionary stages passed through ages ago by the remote ancestors of
Citrus.
3. Pleiospermium
sumatranum Swing. Jour. Arbor. 20:260. 1939. Illus. Swingle,
loc. cit. pl. 3, figs. 3, 4; fig. 3-19 this work.
Type.—Sumatra, Paanus (?) (Korthals, No. 960). Rijks Herb.,
Leiden.
Distribution.—Known only from
Korthals' collections, probably from a single
locality.
Common name.—Sumatra
orangeaster.
Shrub or small tree; twigs
cylindrical, slender (1-2 mm diam.), brownish-gray (in dried herb. spec.),
fruiting twigs spineless, internodes 2-3 cm long; leaf blades broadly
lanceolate, 8-17 cm long, 5-8 cm wide, acuminate at apex, acumen about 9-13 mm
long, often curved slightly to right or left, 3-4.5 mm wide at base, but at the
very tip 1.5-2 mm wide, blunt or slightly emarginate, base cuneate (angle
75°-85°), blade 10-16 X 5-6.5 cm, petiole 18-20 mm long, winged above, 3.5-6.5
mm wide, but wingless for 4-5 mm up from attachment at base, lateral veins
10-12 but of unequal prominence, some strongly elevated above lower surface,
others faint and not prominent, making angles of 60°-75° with the midrib,
margins entire, very faintly undulate, oil glands very numerous, evenly spaced
all over the blade, 0.2-0.5 mm apart, plainly visible below, almost invisible
above, petioles 10-20 mm long, 3-7 mm wide, winged at top, wingless at base;
inflorescences axillary, simple or rarely bifurcate, apparently sometimes
terminal on short lateral twigs; pedicels of nearly ripe fruits 1-1.5 cm long,
2-3 mm wide; flowers unknown; calyx with 5 acute lobes, 1-2 mm wide and 2-2.2
mm long, minutely pubescent below, margins sparingly short-ciliate; fruits
subglobose, 18-20 mm diam., with 3 locules lined with a dense layer of short
pulp-vesicles, 1-2 mm long, 0.4-0.7 mm wide, peel with numerous oil glands
1.5-1.8 mm thick.
This species of
Pleiospermium has remarkably small pulp-vesicles lining the dorsal walls
of the three locules of the fruit. The pulp-vesicles are very
numerous, ovoid, bluntly rounded at the tip, 1 to 2 mm long, 0.4 to 0.7 mm
wide, slightly narrowed at the base but not imbedded in the locule wall,
cortex very thin, composed of indistinct cells, contents sharply delimited
from the cortical layer and soon breaking down into a spongy mass of oily or
resinous (?) matter looking almost exactly like the contents of the oil glands
in the peel of the fruit (see fig. 3-19). Apparently these very small pulp-vesicles
secrete a translucent liquid that completely fills the locules of the
fruit. In dried herbarium specimens this liquid hardens to a dark
reddish-brown, resin-like mass that splinters when struck a sharp blow.
This Sumatran species of Pleiospermium and
its two close relatives from Banguey Island and Borneo, P. longisepalum
and P. latialatum, all have leaves very similar to those of Citrus
except for the more or less acuminate leaf tips.
A fungus, found on the fruits of the type
specimen, was identified by Bitancourt and Jenkins (1938, p. 388) as a species
of Elsinoë much like the scab fungi, E. fawcettii and E.
australis, that attack Citrus.
4. Pleiospermium longisepalum Swing. Jour. Arnold Arbor.
20:259. 1939. Illus. Swingle, loc. cit. pl. 3, fig. 5.
Type.—Banguey Island (off
northeastern point of Borneo) (Castro and Melegrito, No. 1348).
Herb. Natl. Arbor., Washington, D.C. (sheet No.
45800).
Distribution.—Known only from
the type collection and from Castro and Melegrito, No. 1713, from the same
locality.
Common name.—Banguey Island
orangeaster.
A small tree or shrub, fruiting
twigs angular, 2-3 mm diam., straight, glabrous, grayish-green, spineless,
vegetative twigs at first angular then cylindric, 2-3 mm diam., both
vegetative and fruiting twigs with faint striae decurrent from the nodes;
leaves 1-foliolate, leaf blades 7.5-11.5 X 3-4.5 cm, elliptical with acuminate
apices turned slightly to one side, but the actual tip (about 2 mm wide)
abruptly rounded or even slightly retuse, margins entire, lateral veins weak,
8-10 irregular pairs, with irregular reticulations between; petioles 10-14 mm
long, clearly articulated with the leaf blades and with the twigs, winged, 3-5
mm wide at top, tapering into a wingless base constituting about 1/4-1/3 the
total length, wingless basal portion often twisted and apparently pulvinoid;
flowers borne singly in the axils of the leaves but the tip of flowering branch
merging into a racemose inflorescence with the leaves reduced or wanting;
pedicels 20-25 X 1.5-2 mm with several minute bracts borne singly, faintly
striate longitudinally and grayish-green like the twigs; pedicels of sterile
flowers persistent, very slender, 15-20 X 0.75-1 mm; sepals persistent,
gray-green, strap-shaped, 9-12 mm long, 2 mm wide at the base, 0.5-0.8 mm wide
at the apex, which is bluntly rounded or slightly emarginate, straight and
strongly reflexed below the ripening fruit, showing one median nerve on the
inside, margins entire; fruits ovoid, tipped by the umbonate, persistent, very
short style-base, glabrous, gray-green, 20 X 15 mm, with 3 locules, having 1 or
2 ovules in each locule; pulp-vesicles very rudimentary, 4-6 mm long (arising
from the dorsal ovary walls and tapering gradually to a blunt point); peel
thin (1.5-2 mm), gray-green, with numerous, slightly sunken, minute oil glands;
seeds 1 or 2, about 11 X 6 X 4 mm, monoembryonic.
This very curious plant was briefly noted by
Merrill (1926, p. 377), who published Swingle’s observations made on a specimen
of it and his own notes made shortly after its discovery. At first
sight it seemed to be allied to Atalantia because of its unifoliolate
leaves. However, it does not have the well-developed, sessile
pulp-vesicles characteristic of Atalantia but instead has rather
rudimentary, corticate ones somewhat like those found in P.
dubium. Then, too, its winged petioles are unlike any found in
Atalantia, Severinia, or related genera, and are almost exactly
like those of other species of Pleiospermium. The long,
persistent, reflexed ligulate sepals are unique in the orange subfamily,
although the evidently related species, P. latialatum, has similar
sepals, persistent, cuneate, and partially reflexed, but shorter.
The other three species of Pleiospermium have short sepals much like
those of many other members of the subtribe Citrinae.
5. Pleiospermium latialatum
Swing. Jour. Arnold Arbor. 20:261. 1939. Illus. Swingle, loc.
cit. pl. 3, fig. 6.
Type.—North
Borneo (Sabah), Tawao (Elmer, No. 21542). Herb. Arnold Arbor., Harv.
Univ., Cambridge.
Distribution.—Known
only from the original collection.
Common
name.—North Borneo orangeaster.
A small
tree or shrub, ultimate twigs slender, 1-2 mm diam., at first slightly angled,
soon terete, very finely puberulent, more or less longitudinally wrinkled,
nodes 1-1.8 cm long; spines lacking (at least on fruiting branch); leaves
1-foliolate, glabrous, shiny above, dull below, oblong-elliptical or
lanceolate, acuminate-caudate at apex, end of acumen linear for 4-6 mm and
1.5-2 mm wide, ending abruptly with an emarginate tip, broadly rounded or very
bluntly cuneate at the base; leaf blade 9-15 X 3.7-7 cm, margins entire,
lateral veins 10-13 pairs, arising at an angle of 70°-75° with the midrib,
curving slightly toward leaf apex, oil glands small, very numerous; petioles
(all detached) short, broadly winged, obcordate, 1.6-2.4 X 1-2 cm. narrowed
into a pulvinoid and minutely wrinkled terete base, 1-2 mm long, 1.25 mm wide
with a narrow channel above formed by the decurrent wings; flowers, not seen,
arising singly (?) at the tips of short axillary branches which bear leaves or
bracts below; calyx puberulent without, sometimes persisting under young
fruits, lobes narrowly cuneate or somewhat rounded, apparently irregular in
size, acute or rounded, 2-4 mm long, 2-5 mm wide, diverging merely at right
angles to the pedicel under the young fruit; young fruits subglobose, 10-14 mm
diam., glabrous, peel green, with numerous oil glands, slightly raised above the
surface with a small central depressed (crateriform); locules 3; pulp-vesicles
conical, very slender, 3-4.5 mm long, acutely pointed, attached to the dorsal
wall of the locule into which their bases are somewhat sunken, cortication very
thin with indistinct cells, contents not degenerating to an oily or resinous
mass as in P. dubium but showing axile droplets of oily substance; seeds
2 or 3, large and plump.
This remarkable
species is known from a single collection from Tawau, in North Borneo (about
175 km southwest of Shibutu Island in Sulu Archipelago, Philippine
Islands). It is obviously related to P. longisepalum but
differs strikingly in its very broadly winged petioles, which are also
relatively short, giving them a very pronounced obcordate outline.
The sepals are sometimes (perhaps always) persistent under the developing
fruit, but are only about half as long as those of P. longisepalum and
are not completely reflexed, but stand out approximately at right angles to the
general direction of the pedicel. Only young fruits were available
for study but they show clearly that the pulp-vesicles of this species, much
like those of P. longisepalum, are very numerous, slender-conical in
shape, and slightly immersed in the dorsal wall of the locule. The
cortical layer of the pulp-vesicle is present but not strongly developed and
apparently sometimes this cortex is so weak that the upper portion of the
pulp-vesicle splits open as the fruit develops. This species has
leaves that strikingly resemble those of Citrus in general
appearance.
XVI. Burkillanthus
Swing.
XVI. Burkillanthus
Swing. Jour. Arnold Arbor. 20:255.
1939.
Type species.—Citrus
malaccensis Ridl. = Burkillanthus malaccensis (Ridl.) Swing.; from
Malacca, Malay Peninsula.
Distribution.—Malay Peninsula: Malacca; northern Sumatra:
Asahan.
Common name.—Burkillanthus or
Malay ghost-lime.
Leaves 3-, 2,- or
1-foliolate; leaflets thin, with numerous strongly marked, subparallel, lateral
veins; petioles variable in length, articulated, narrowly winged or sometimes
merely margined, with pulvinoid bases; petiolules very short, pulvinoid;
inflorescences, few-flowered clusters in the leaf axils; flowers large, 5-merous
with 10 stamens; pedicels short, about the length of the styles; calyx deeply
5-lobed; petals 5, spathulate; ovary obclavate, minutely and sparsely
pubescent, merging abruptly into the much narrower style, which is nearly as
long as the ovary, with 5 locules, each with 22-26 ovules arranged in 2 rows;
fruits very large, ovoid, more than twice as long as the petioles, with a thin
peel dotted with numerous oil glands covering a thin, hard, woody endocarp;
pulp-vesicles sessile (not deeply counter-sunk) on the dorsal locule walls,
very long, swollen near the base, then cylindrical, then tapering to an acute
apex, corticate, with translucent inner core not breaking down into an oily or
resinous (?) mass; seeds very large, with a thin, slightly wrinkled testa,
immersed in mucilaginous gum and mingled with the collapsed rudiments of the
pulp-vesicles; embryo single with light buff (not green) cotyledons.
This strange plant seems to be an evolutionary
freak of nature perhaps descended from some ancestral form more or less like a
trifoliolate Pleiospermium such as P. alatum or a uni-, di-, or
trifoliolate species like P. dubium.
The pulp-vesicles of Burkillanthus are stalkless and somewhat resemble
those of Pleiospermium; however, they are very much larger and less
obviously corticate, with a central core which does not disintegrate into an
oily or resinous (?) mass resembling that of an oil gland. These
pulp-vesicles of Burkillanthus seem to represent a very interesting
developmental stage about midway between the pulp-vesicles of
Pleiospermium and those of Clymenia, one of the True Citrus Fruit
Trees.
It is
remotely possible that the curious, cavernous lining of the radial locule
walls in Swinglea may, upon study of fresh material, prove to be composed
of still more primitive structures analogous to the pulp-vesicles of
Burkillanthus. the [sic] pistil of B. malaccensis
(see fig.
3-20) is extraordinarily like that of Swinglea glutinosa in size,
shape, number, and arrangement of the ovules, et cetera, except that in
Swinglea the ovary is hypermerous, having 8 to 10 locules (not five as in
Burkillanthus) and oil glands occur in all parts of the pistil, including
the stigma (see fig. 3-20). However, the fruits of Swinglea
have a very different gross structure from those of Burkillanthus and the
seeds of Swinglea are not smooth but are covered with flattened laciniate
paleae.
Cockrell (1935) found that the wood of
B. malaccensis, as it grows near Asahan, Sumatra, is almost exactly like
that of Micromelum pubescens (one of the Very Remote Citroid Fruit
Trees), differing from it only slightly in texture and not showing any
"'reasonably constant basis for separation." However, the wood of
Burkillanthus differs decidedly from the wood of all the other genera of
rutaceous plants, 16 in all, that he studied, including eight genera of the
orange subfamily, viz.: Glycosmis, Clausena, Murraya,
Merrillia, Feronia, Aegle, Atalantia, and
Citrus. He did not, however, have wood of
Pleiospermium or of Swinglea to compare with that of
Burkillanthus.
Burkillanthus
malaccensis (Ridl.) Swing. Jour. Arnold Arbor. 20:257. 1939.
Citrus malaccensis Ridl. Fl. Malay Penin. 1:359. 1922.
Illus. Swingle, loc. cit. pls. 2 (1/2 nat. size, not nat.
size) and 3 (figs. 1, 2); figs. 3-20 and 3-21 this work.
Type.—Malay Peninsula, Malacca Territory, Nyalas (Goodenough, No.
1273). Cotype: Malacca State, Bukit Sedanan (Derry, No.
1106). Both in Herb. Bot. Gard.,
Singapore.
Distribution.—Malay
Peninsula: Malacca; Sumatra: Asahan.
Common name.—Burkillanthus or Malay ghost-lime.
A tree up to 14 m high, some branches with
stout, straight, usually paired spines, others spineless, with leaves mostly 1-
or 2-foliolate, rarely 3-foliolate; leaflets subcoriaceous but thin, broadly
lanceolate or elliptic-lanceolate, acute or more or less acuminate at the tips,
which are blunt at the very apex, terminal leaflets and 1-foliolate leaves
variable in size, often large, 12.5-27 X 4.5-11.5 cm, lateral leaflets much
smaller, 6.5-8 X 3-4 cm, with short, pulvinoid petiolules 2-2.5 mm long, lateral
nerves (of larger leaflets) about 12-18 on each side arising at a very wide
angle (70°-85°) with the midrib, running subparallel and only slightly curved
for 2/3 or 3/4 the distance from the midrib to the margin, then branching
dichotomously, base broadly cuneate, articulated with the petiole by means of a
pulvinoid petiolule, margin subentire, very slightly coarse-crenate; petioles
of the 2- or 3-foliolate leaves 4-5 cm long, often narrowly winged (3-6 mm in
total width) at apex, narrowing toward the base, often merely margined even at
the apex; petioles of 1-foliolate leaves usually shorter, 1-3, rarely 4, cm
long, sometimes almost wingless when short; inflorescences few- (usually 1-4-)
flowered, axillary clusters, flowers large, 5-6 cm diam., borne on pedicels 5-7
mm long; calyx of 5 lanceolate-acuminate, acute sepals with large oil glands,
free nearly to the base; petals 5, subspathulate, with oil glands in the
broadened upper part, 20-25 mm long, 3.5-4.5 mm wide, tapering into a long
slender base 1-1.5 mm wide; stamens 10, filaments free, long, slender, anthers
rather small, oblong; ovary obclavate, 6.5-7 mm long, 2.5 mm diam. at the
broadest part (about 2/3 the distance from the top of the disk to the base of
the style, at a point shortly below the tips of the locules) and 1.8-2 mm diam.
at the base, with 5 locales, each with 22-26 ovules in 2 rows in the upper 2/3
of the locular cavity (see fig. 3-20), the entire ovary covered with short sparse pubescence
and with a few small oil glands dotted over the upper half only; style slender,
5.5-6 X 0.5-0.7 mm, with small oil glands, glabrous, merging abruptly into the
tip of the ovary without any definite demarcation, swelling gradually at the
tip and surmounted by a depressed globose stigma, 1 mm high, 1.6 mm wide,
without oil glands; disk short, cylindrical, 0.6-0.7 mm high, 1.4-1.5 mm wide;
fruits large, ovoid, 10-11 X 8-9 cm, outer peel thin (2-3 mm), roughened with
numerous slightly depressed oil glands, fitting closely on woody endocarp, 2-3
mm thick, very hard in dried fruits, that shows on its outer surface numerous
slightly depressed areas 1.5-3 mm diam.; pulp-vesicles sessile, 15-33 mm long,
with bases 2.5-4 mm diam., tapering into long cylinders, 1.5-2 mm diam., which
often end in very acute tips; corticate with small tangentially compressed
peripheral cells and larger, thinner-walled central cells somewhat elongated in
their radial diameter; locules of the ripe fruit almost filled with large
seeds, apparently imbedded in mucilaginous gum, perhaps secreted by the
pulp-vesicles; seeds broadly obovate, tapering to an acute point at the base
where the seed is attached, 22-27 X 16-18 X 7-10 mm, smooth, cream-colored, with
a cinnamon brown chalazal cap covering the broad end of the seed for 6-8 mm,
monoembryonic with pale buff (not green) cotyledons, testa thin, papery,
irregularly wrinkled on dried seeds.
Burkill
published (1931, p. 218) very interesting notes on this species, as follows:
"The characters of this extremely interesting plant have not been brought into
adequate prominence. It was collected in the year 1893 twice in the
interior of the Territory of Malacca, and is represented in the herbarium of
the Botanic Gardens, Singapore, by a sheet of each gathering. Derry
obtained it with flowers in January, and Goodenough with fruit in
July. Derry's locality is given as Bukit Sedanan; Goodenough's as
Nyalas; these places are 6-8 miles apart. Upon the two sheets there
are ten leaves; and upon each sheet one is compounded by having a single
lateral leaflet. Other leaves have a scar at the apex of the
narrowly winged petiole and therefore count as compound; others have no scar,
and obviously are simple. Trifoliate leaves do not exist; but it
would be curious if they are never produced. There is a conspicuous
joint between the petiole and the leaf-blade or blades. The flower
suggests that of Merrillia caloxylon Swingle. The sepals are
in shape as those of Atalantia and have large glands. The
petals are nearly spathulate, and with glands in the broadened upper
part. The stamens apparently are 10, free, with long filaments and
rather short anthers. The ovary is hairy and fluted with five
grooves; it is hard to distinguish the exact place where the gynophore passes
into it. The style is as long as the ovary and ends in a large
stigma. There are numerous ovules. For other points
the original description may be consulted."
This remarkable plant was first collected in flower by R. Derry (at that time in
charge of the Malacca forests) at Bukit Sedanan, near Malacca, in January,
1893. On July 9 of the same year, J. Goodenough collected at
Nyalas, only some eight miles distant from Bukit Sedanan, a specimen with ripe
or nearly ripe fruits (see fig. 3-21) (fide Burkill, in letter dated January 13,
1937). Thanks to the kindness of R. E. Holttum, then director of
the Botanic Gardens at Singapore, Swingle was privileged to have the loan of
the two type specimens noted above.
E. D.
Merrill directed attention in June, 1938, to a tree which he had assigned to the
subfamily Aurantioideae, collected by B. A. Krukoff (No. 4233) in
November-December, 1932, in the Nassihi Forest Reserve near Asahan in northern
Sumatra (about 335 km west-northwest of Malacca). Herbarium
specimens kindly lent by the New York Botanical Garden and the Arnold Arboretum
showed that this plant was almost identical with the type specimens of
Citrus malaccensis Ridl. The twigs of Krukoff's specimens
were in part spineless but in part armed with stiff, stout, straight, abruptly
pointed, usually paired thorns 2 to 3 cm long, and 2 to 3 mm in diameter at the
base. The leaves were uni-, di-, or trifoliolate. A
giant unifoliolate leaf in the New York Botanical Garden herbarium had a
petiole 2 cm long and a broadly lanceolate blade, 27 by 11.5 cm,
short-acuminate at the apex, cuneate at the base. It had 17 or 18
principal lateral veins, and about 9 or 10 shorter laterals, arising at angles
of 70° to 80° with the midrib (see Swingle, 1939a, p. 262, pl.
2). The ovoid fruit measured about 11 cm long and 9 cm
wide. The inner side of the dorsal walls of the locules were
thickly covered with cylindrical or subclavate (?) saclike pulp-vesicles, as
described above. No similar pulp-vesicles are known in any other
genus of citrus fruit yet studied.
In a letter
to Swingle, dated April 19, 1940, R. E. Holttum reported that a collector whom
he had sent to Bukit Sedanan, the type locality of B. malaccensis, found
this species growing there in quantity "in primitive forest, but only on the
ridges, where the shade is less than on the valley slopes." The
trees were about 30 feet (9.15 m) high and bore half-grown fruits.
In June, 1941, a large fruit, slightly over 10
by 16 cm, was sent to Swingle by air express from the Singapore Botanic
Gardens. It reached Washington in good condition, which made it
possible to study fully developed pulp-vesicles in the living
state. This fruit contained more than 120 very large, mature
seeds. (See fig. 3-21.)
This remarkable
tree should be studied in the living state and possibly tested as a rootstock
for Citrus. It is called by the Malays limnan hantu
("ghost lime"), a name also applied to wild forms of Citrus according to
Burkill (1931, p. 221), perhaps because its large fruits are not unlike those of
some species of Citrus in size and shape.
XVII. Limnocitrus
Swing.
XVII. Limnocitrus
Swing. Jour. Arnold Arbor. 21:2.
1940.
Type species.—Paramignya ?
littoralis Miq. = Limnocitrus littoralis (Miq.)
Swing.
Distribution.—Java: Bali; South
Vietnam.
Common name.—Limnocitrus or
swamp-orange.
A shrub with glabrous twigs
having stout, axillary, single spines at the nodes; leaves simple, coriaceous,
glabrous, broad-elliptical or obovate, bluntly pointed at the tip and broadly
cuneate at the base; petioles short (less than 1/10 as long as the leaf blade),
pulvinoid, not articulated with the leaf blade; inflorescences terminal, short,
corymbose; flowers 4- or 5-merous, borne on very short, glabrous pedicels;
flower buds oblong, greenish; calyx 4-5-lobed, lobes triangular, obtuse; petals
linear-lanceolate, obtuse, white; stamens 10, filaments free, glabrous, anthers
linear; disk annular; ovary oblong-ovate, with 15-20 narrow hirsute ridges,
with a single large oil gland over each of the 4 or 5 locules, each having 2
ovules; style slender, sparingly hirsute, stigma slightly capitate, distended
with large oil glands between the stylar canals; fruit globose or subglobose,
rather large (3.5-4 cm diam.), with a gland-dotted peel, becoming orange-colored
when ripe, filled with very numerous, slender, fusiform pulp-vesicles, slender
and acutely pointed at tip and abruptly truncate at the broad base about
1/3-2/3 as wide as the broadest part, which is near the base, with a slender
axile [sic] column of whitish granules (oil or resin drops?) in the
center; seeds large, flattened-oval, with a hard, creamy-white testa,
monoembryonic, embroyo green, cotyledons dotted with oil glands.
This remarkable plant has been placed in several
different genera by the few botanists who have discussed it. It has
certain resemblances to the species of Pleiospermium but differs from
them in several important characters: (1) in having simple leaves with very
short pulvinoid petioles; (2) in having slender, fusiform pulp-vesicles with
very slender, acute tips and slightly narrowed at the base where attached; (3)
in having thick, sparingly veiny leaves; (4) in having straight, very stout,
single spines in the leaf axils.
Limnocitrus is distinguished from Atalantia by its very
slender-tipped pulp-vesicles, which are not conical and do not have broad bases
deeply immersed in the locule wall, as in Atalantia, but are fusiform and
narrowed into a definite, though very short, basal stipe. The leaves
are thicker and differently veined and the whole plant has a different facies
from that of Atalantia.
In many ways
the pulp-vesicles of Limnocitrus are intermediate between those of the
True Citrus Fruit Trees (Citrus and the five most closely related genera)
and those of the Near-Citrus Fruit Trees (Citropsis and Atalantia)
and some of the Primitive Citrus Fruit Trees, especially
Pleiospermium. In most of its other characters,
Limnocitrus stands very much alone. The leaves are different
in texture, venation, and general appearance from those of all the other
twelve genera in the subtribe Citrinae. However, similar, although
not identical, leaf and twig characters are found in Merope and
Pamburus, two monotypic genera of the Minor Citroid Fruit Trees, subtribe
Triphasiinae, which also grow in brackish
marshes.
Limnocitrus littoralis (Miq.)
Swing. Jour. Arnold Arbor. 21:3. 1940. Paramignya ?
littoralis Miq. Ann. Mus. Bot. Lugd.-Bat., 1:211. 1863, and Teijsmann
& Binnendijk, Natuur. Tijdschr. Nederl.-Indie 27:41. 1864; Limonia
littoralis (Miq.) Backer, 1911; Atalantia littoralis (Miq.) Guill.
1913; Pleiospermium littoralis (Miq.) Tan. Bul. Mus. Hist. Nat. Paris, 2
ser. 2:162. 1930. Illus. Valeton, Icon. Bogor. 4:163, pl.
349. 1912; Swingle, loc. cit. pl. 1; fig. 3-22 this work.
Type.—Java, Rembang, on seashore (Teijsmann). Rijks. Herb.,
Leiden.
Distribution.—Java: Bali; South
Vietnam: Annam, Cochin China.
Common
name.—Limnocitrus or swamp-orange.
This
species was based on a specimen collected by Teijsmann on the seashore at
Rembang on the north coast of Java. Miquel's original description
reads, in translation, as follows: "Spines solitary, straight, arising nearly
at right angles to the twig; leaves simple, with very short petioles, acute or
acutish at the base, broad- or obovate-elliptical, apex obtuse or somewhat
protracted-obtuse and emarginate, coriaceous, pellucid-punctate, reticulate,
venose below, [margins] very slightly crenulate; leaves 2-3 in. [5-7.6 cm]
long; flowers in terminal tufts, forming much contracted cymes, with very
short, thick pedicels; calyx unequally 5-toothed; petals 5, elliptic-oblong,
imbricate; stamens 8, filaments very short, anthers linear, marked with an
oblong gland on the back, joined together in the bud; ovary inserted on a short
disk, 5-angled, sparsely pilose, 5 locular, ovules 2 in each locule,
subcollateral; style thick, stigma slightly capitate, obscurely 5-lobed; fruits
globose, 2- or 3-seeded, about the size of apricots, with distinct, empty
locules."
Another description published almost
simultaneously by Teijsmann and Binnendijk (doubtless based in part on
independent observations of the species by Teijsmann, who discovered it at
Rembang) supplements Miquel's original description. Omitting
obvious duplications, this reads, in translation, as follows: "Branches
cinereous, twigs very glabrous; spines 1/2 in. [12.5 mm] long, stout; leaves
emarginate at the apex, rarely obtuse; racemes [sic] short, sessile…calyx
4-5-lobed; filaments 10, free; ovary pubescent, 4-5-locular, ovules paired;
fruits baccate, very fetid, 2.3 cm diam., locules by abortion 1-seeded."
In the notes following the technical description
they add, regarding the fruit, that it is "depressed-globose, about the size of
a lime, becoming orange-colored…4-5-locular…pulpy; rind,
leathery-glandular. Seeds large, testa membranaceous,
white. Cotyledons fleshy, plano-convex, green."
One of the most important characters of the
species, overlooked by Miquel but brought to light by Teijsmann and Binnendijk,
is that the locules of the fruit contain pulp. This was mentioned a
few years later by Kurz, who lived for some years in Java, where he doubtless
saw Teijsmann's specimens collected at Rembang. Kurz wrote (1875,
p. 136, footnote): "The berries of P[aramignya] littoralis
Miq., a species closely allied to P. angulata [= Merope
angulata], have pulp, but the dried ones appear pulpless."
Fortunately Swingle was able to supplement the
above-quoted descriptions and a later one by Valeton (l.c., p. 163), all based
on a single collection made at Rembang, Java, from excellent material from
Indo-China. Flowering specimens studied were from plants growing in
a hedgerow near houses at Tourane, Annam (coll. by J. and M. S. Clemens, No.
3263), in Herb. Arnold Arboretum; fruiting material was from Nhatrang, Annam
(Aug. Chevalier, No. 30526, February 6, 1916), in Herb. National
Arboretum. This material showed the following characters:
A shrub or small tree 2 m high with stout,
straight, single spines; leaves simple, glabrous, broadly oval, bluntly pointed
at both ends, margins entire or faintly crenulate; leaf blades 5-7.5 X 3-4 cm,
with 8-11 pairs of lateral veins, arising at angles of 40°-60° with the midrib,
usually straight but sometimes bending to the right or left to unite with the
neighboring lateral vein, tertiary or quaternary veinlets forming coarse,
irregular, inconspicuous reticulations, oil glands small, very numerous;
petioles cylindrical, 3.5-7 X 1-1.5 mm, pulvinoid, minutely puberulous on the
upper side inside the shallow channel bounded by the decurrent leaf margins;
inflorescences terminal (sometimes also small ones in axils of last few
leaves), corymbose, 3-4 X 2.5-5 cm, pubescent, peduncles abundantly and
divaricately branched, divided into nodes (5-10 X 1.5-2 mm) by thick,
blunt-pointed, pubescent, caducous bracts, 1-5 X 0.8 mm, pedicels 2.5-3.5 mm
long, expanding gradually into the base of the calyx without any articulation;
calyx glabrous without, 4-5-lobed, lobes bluntly pointed, finely pubescent
within, with short-ciliate margins; petals white within, green without in the
bud and greenish-white when expanded, linear-oblong, blunt-tipped, 10-11 X 3
mm, glabrous except for scattered puberulence near the tip; stamens with
glabrous filaments, anthers long and slender, 4.5 X 1 mm, with a large dorsal
oil gland; disk cup-shaped, 0.5-0.6 mm high, 1.5-1.6 mm wide, enclosing loosely
the base of the ovary for 0.2-0.25 mm; pistil 10-11 mm long; ovary obovoid,
4.5-5 mm high, 1.7-2 mm wide, flattened at top, obscurely ribbed in middle
portion with 15-20 slight ridges from which arise scattered long hairs,
pentagonal at the top because of large sunken oil glands (1 over each locule),
locule [sic] 4-5, each with 2 pendant collateral ovules; style 6 mm
long, 1.8-2 mm thick, terete, sparingly pilose above, increasingly so toward the
base, stigma depressed capitate, 1.8-2 mm wide, 0.7-0.8 mm high, with 4-5
radially arranged, linear stylar canals, with 2 (rarely 1) large oil glands
between each adjoining pair of stylar canals; fruits subglobose, 37-40 mm
diam., peel thin, dotted with oil glands, containing numerous, very slender,
fusiform pulp-vesicles, 9-11 mm long, gradually tapering to 0.5-0.8 mm diam. at
the base where attached to dorsal wall of locule, the broadest portion (1.6-1.8
mm) of the pulp-vesicle is situated about 1/10-1/6 of the distance from the
base to the tip, the slender apical portions often matted and tangled in dried
specimens; seeds long obovate, 17-18 X 9-11 X 3.5 mm, with a smooth, firm,
yellowish-gray testa, monoembryonic; cotyledons green, gland-dotted.
This little-known species apparently has a fairly
wide distribution, as it has been found in Annam and Cochin China in South
Vietnam, as well as in Java and Bali. However, it has been
collected only a few times since it was discovered by Teijsmann some
seventy-five years ago at Rembang in Java, where apparently it has not been
seen since.
It has, however, been collected
several times near Nhatrang in Annam (see fig. 3-22), and once at Tourane. It grows in tidal
swamps subject to overflow with brackish water at spring tides.
Guillaumin (1913, p. 442), in describing the plant as found in the vicinity of
Nhatrang, wrote: "The fruits, as a matter of fact, resemble small oranges
about 4 cm in diameter, with five locules almost completely occupied by two
very large seeds surrounded by pulp formed by succulent hairs, as in the genus
Citrus and in Atalantia citroides." He went on to say
that it is called kim-do-um by the natives at Nhatrang and that, since it
grows on sea beaches, it is probably
salt-resistant.
Limnocitrus, from its
habitat, is almost certain to possess a high tolerance to salinity in the soil
moisture. Citrus can be grafted on Merope, but has made only unsatisfactory
growth. It is probable that Citrus would grow better when
grafted on Limnocitrus, to which it is much more closely related than to
Merope (a member of another subtribe). Such a rootstock might
be expected to be useful wherever Citrus is grown in soils showing a high
salinity in the soil moisture, as has been found to be true with the xerophytic
Eremocitrus, native to more or less saline flats in the semiarid plains
of Australia.
XVIII. Hesperethusa
Roem.
XVIII. Hesperethusa
Roem. Syn. Hesper. 1:31. 1846. Limonia Sec. III, Hesperethusa
Engl. Die Nat. Pflanzenfam. 3(4):190.
1896.
Type species
(monotype.—Limonia crenulata Roxb. = Hesperethusa crenulata
(Roxb.) Roem.; from British India.
Distribution.—Northern India, Burma, southwestern China, Thailand, South
Vietnam, Laos, Cambodia.
Common
name.—Hesperethusa.
Slender, thorny trees
or shrubs; leaves persistent, odd-pinnate; petioles and rachis broadly winged
(petioles sometimes more narrowly winged); inflorescences axillary,
few-flowered; flowers small, 4-merous (except stamens, twice as many as
petals); ovary with 4 locules, 1 ovule in each locule; fruits small, 1 or more
of the 4 segments with a single seed, surrounded by scanty, bitterish pulp
containing large rudimentary sessile pulp-vesicles of irregular size and shape;
seeds hard, smooth, germinating with epigeous
cotyledons.
Hesperethusa has apparently
no near relatives, differing widely from all the other genera of the Citrus
Fruit Trees except Citropsis in having odd-pinnate leaves with three to
four pairs of small, opposite, crenate leaflets and broadly winged petioles and
rachis segments. Pinnate leaves were undoubtedly borne by the
remote ancestors of all the Citrus and Citroid Fruit Trees (the tribe
Citreae). The characters persist today only in two out of the
thirteen genera (Hesperethusa and Citropsis)of the second
subtribe, Citrinae, and in two out of the seven genera (Feronia and
Feroniella) of the third subtribe, Balsamocitrinae. None of
the eight genera in the first subtribe, Triphasiinae, have pinnate
leaves. In other words, only two of the three subtribes of the
tribe Citreae have genera with pinnate leaves, and out of a total of
twenty-eight genera in the tribe only four have pinnate
leaves.
Hesperethusa has some other
points of similarity with the African genus Citropsis aside from pinnate
leaves. The seeds of both genera have a hard testa, cream-yellow in
color, composed of radial prismatic cells, some with very thick
walls. The little known Citropsis daweana from Portuguese East Africa has the smallest
leaflets of any of the species of Citropsis, with widely winged rachis
segments but nearly wingless petioles, making the leaves look somewhat like
those of certain forms of Hesperethusa crenulata.
The very small fruits of this monotypic genus
show rudimentary pulp-vesicles attached to the dorsal wall and apparently also
to the base of the locule. There are relatively few of these
pulp-vesicles in each locule of the developing fruit; they are not clearly
differentiated into basal and apical portions but are irregularly rounded and
seem to be thin-walled; they vary greatly in size, and are often polygonal from
mutual pressure. These rudimentary pulp-vesicles can be seen in the
sections12 of the fruits of paratypic
material in the Kew Herbarium which was collected by Roxburgh in the last
quarter of the eighteenth century and from which he drew up the description of
Limonia crenulata, the type species of the genus.
The pulp-vesicles of Hesperethusa that
arise from the dorsal wall of the locule appear like small vesicles adhering to
the wall by a broad base and are broader than high. The few
pulp-vesicles that arise from the base of the locule are more or less slender
and elongated but do not have any clearly marked stalk. The shapes
taken by the pulp-vesicles seem to be governed by the free space available into
which they can expand as they develop. The pulp-vesicles attached
to the dorsal wall are somewhat like those of Pleiospermium in shape but
they do not show any definite cortical layer and furthermore do not show any
degenerations of the central portion into an oily or resinous (?)
mass.
Hesperethusa can be grafted on
Citrus and such grafts live and grow for many years.
Dr. Anna E. Jenkins (1936, pp. 71, 73) discovered
on the fruits of Hesperethusa from India a fungus, Elsinoë sp.,
very like the scab fungi, E. fawcettii and E. australis, that
attack species of Citrus.
Hesperethusa crenulata (Roxb.) Roem. Syn. Hesper. 1:38. 1846.
Limonia crenulata Roxb. Pl. Corom. 1795; L. acidissima Auct. (non
L.). Illus. Rheede & Draakestein, Hort. Malabar. 4:31, pl. 14.
1679; Roxburgh, loc. cit. 1:pl. 86; Talbot, Forest Fl. Bomb.
Presid. 198, fig. 121. 1909; Engler, Die Nat. Pflanzenfam. 3(4):190,
figs. J, K, L. 1896; Swingle, in Bailey, Stand. Cycl. Hort.
3:1478, fig. 1825. 1915; fig. 3-23 this work.
Type.—Peninsular British India (Roxburgh). Herb. Brit. Mus.,
London (fide Tanaka).
Distribution.—West Pakistan, India, Burma, Thailand, southwestern China,
South Vietnam, Laos, Cambodia.
Common
name.—Hesperethusa.
Talbot (1909, pp. 198,
199) described this species as follows: "A spinous, glabrous, small tree:
spines straight 0.5"-1" [12-25 mm] long. Leaves imparipinnate;
leaflets 2-3 pairs, 1"-2" [2.5-5.1 cm] by 0.5"-1" [1.2-2.5 cm], conspicuously
gland-dotted, sessile, ovate, emarginate, crenulate; terminal leaflet usually
largest; rachis and petiole broadly winged, jointed; joints of rachis
obovate. Racemes short, axillary, pubescent, subumbellate,
few-flowered. Flowers 4-merous. Calyx small, glandular,
4-lobed; lobes ovate. Petals 4, fragrant, white, 0.25" [6 mm] long,
elliptic. Disk annular or stipitate. Ovary 4-celled
with 1 pendulous ovule in each cell; style short. Fruit globose,
1-4 seeded, 0.25" [6 mm] in diameter, black when ripe; pedicels 0.75" [19 mm]
long."
Hesperethusa has a wide range
from West Pakistan to Burma, southwestern China on to southern Indo-China
(Cambodia). In West Pakistan and India, it occurs commonly in the
sub-Himalayan tract from Ravi River (Long. 75° 30' E.) eastward, ascending to
4,000 feet [1,220 m] altitude.
This species is
a small tree reaching a height of 25 to 30 feet (7.6 to 9.14 m); it grows
commonly on dry hills or in dry jungles. It was said by Haines
(1921, vol. 2, p. 163) to be "subdeciduous at the time of
flowering." The wood is largely used in Bihar and Orissa, India, for
cart axles. Several authors have stated that the spines are
sometimes paired and that the leaves often have four pairs of
leaflets. The inflorescences often bear from one to several foliage
leaves at the base, and leaf spurs (Kurztrieben), with very short
internodes, are often borne in the axils of the leaves of older
twigs. The petioles of the leaves on such Kurztrieben are
often more narrowly winged than the rachis segments of the same leaves and are
sometimes almost wingless. The fruits are said by many authors to
be very acid, but Haines (1921, vol. 2, p. 163) reported them to be "intensely
bitter (not acid)."13 Guillaumin (1911,
p. 660) spoke of the seeds being "immersed in a mucilage," but no one seems to
have realized up to now that the locules of the fruit contain rudimentary
pulp-vesicles. Hesperethusa crenulata produces a hard,
close-grained, light yellow wood in India, and its leaves, fruits, and roots
are used in making medicine in that country. According to Watt
(1890, vol. 4, p. 642) the fruit is exported from India "to the Arabian coasts,
where it is used as a condiment with fish, meat, etc., being powdered along
with the spices commonly used in cooking."
Hesperethusa crenulata is promising as an ornamental because of its
beautiful, feathery, green foliage (see fig. 3-23). Grafts of it on Citrus were still
growing in the greenhouses of the Agricultural Research Service at Washington
after 20 years, although the union of the stock and scion was somewhat
swollen. It can also be grafted on Swinglea glutinosa (the
Philippine tabog), which belongs to the subtribe Balsamocitrinae.
Group B. THE NEAR-CITRUS FRUIT
TREES
This group includes two genera,
Citropsis and Atalantia, that show well-developed pulp-vesicles
having broad sessile bases and conical sides that taper regularly to the acute
apices.14 These pulp-vesicles are arranged more
or less radially with the bases at the periphery of the fruit locules attached
to the dorsal walls of the locules and imbedded in the inner layer of the
rind. The conical pulp-vesicles usually point toward the center of
the fruit unless deflected by the seeds, which (if present) are attached at the
inner angle of each locule. The genus Citropsis, native to
tropical and subtropical Africa, has eleven species, all but one with pinnate
or trifoliolate leaves resembling in several important characters those of the
genus Hesperethusa of group A; Atalantia, native to southeastern
Asia (including Ceylon), likewise has eleven species, but all have unifoliolate
or simple leaves much resembling those of Citrus of group C in size and
shape but differing in their more leathery texture and finer venation.
Both Citropsis and Atalantia are
obviously related to the True Citrus Fruit Trees of group C, and some three
species of Citropsis and two or three special of Atalantia (all
that have been tested) can be grafted on Citrus, and vice
versa. These two genera are intermediate in character between the
Primitive Citrus Fruit Trees of group A and the True Citrus Fruit Trees of
group C; like the genera in group A they contain many obvious connecting links
between Citrus and the Minor Citroid Fruit Trees of the subtribe
Triphasiinae.
XIX. Citropsis (Engl.)
Swing. & M. Kell.
XIX.
Citropsis (Engl.) Swing. & M. Kell. Jour. Agr. Res. 1:421. 1914.
Limonia Sec. citropsis Engler, Die Nat. Pflanzenfam.
3(4):189. 1896.
Type
species.—Limonia preussi Engl. (from Elephant Lake, Cameroons) =
Citropsis articulata (Willd.) Swing. & M. Kell. (ranging from Togo to
Cameroons and to Belgian Congo).
Distribution.—Not uncommon from Uganda to Nigeria and Sierra Leone, and
south to Republic of the Congo, Angola, and
Mozambique.
Common name.—African
cherry-oranges.
A genus of Near-Citrus Fruit
Trees found in tropical Africa. Shrubs or small trees with 1 or 2
well-developed spines (C. angolensis is said to be spineless) in the
axils of the leaves; leaves pinnate, sometimes 3-foliate [sic], rarely
1-foliolate (simple in C. tanakae); rachis and petiole usually broadly
winged (wingless in C. le-testui), flowers 4- or 5-merous, usually in
dense axillary clusters or short racemes; stamens twice as many as the petals,
filaments free, flattened laterally and in some species cohering laterally at
the base in groups; ovary with 4 or 5 locules, each with a single ovule; fruit
spherical, small (2-3 cm diam.), with pulp-vesicles broad at the base where
they are attached and tapering to an acute tip; seeds large (10 X 6 X 4 mm);
cotyledons hypogeous in germination.
This
genus is certainly closely related to Citrus, in spite of the fact that
all but one species of Citropsis have pinnate or trifoliolate
leaves. It belongs, along with Atalantia, to group B, the
Near-Citrus Fruit Trees, of the subtribe Citrinae. Both of these
genera have small, subglobose fruits, 1.5 to 3 cm in diameter, with 3 to 5
locules, in which occur broad-based, sessile, conical, radially arranged
pulp-vesicles. All the species of Citropsis have leaves, or
leaflets, very similar to those of Citrus in shape, venation, and
texture, whereas the leaves of the species of Atalantia are thick, more
leathery, and much more veiny than those of Citrus.
The seeds of Citropsis are plump and
broadly ovoid with a very hard, smooth testa. The hilum is an oval
or ovate aperture bordered by the incurved testa, which is here very hard and
thick. No other genus of the subtribe Citrinae has seeds as plump
and as hard-shelled. The flowers of Citropsis differ greatly
in the various species with respect to size and proportion of parts, and
especially with respect to the morphology of the pistil. The leaves
differ even more than the flowers in the different species, but fortunately for
the taxonomist the leaf and flower characters are not closely correlated; that
is, a pair of species having very similar leaves, like C. schweinfurthii
and C. angolensis, may show easily recognized differences in the
flowers. Citropsis schweinfurthii and C. gilletiana
have similar flowers, with only slight differences in the pistil, but very
different leaves.
The species of
Citropsis look like missing links, i.e., surviving forms of the remote
ancestors of Citrus and may, in fact, be such forms that migrated tens of
millions of years ago from the southeastern Asiatic homeland of the tribe
Citreae and found safe and congenial refuge in the tropical forests of Africa,
where they have persisted with little change ever since. Once
safely in Africa, the ancestors of Citropsis were freed from
competition with the numerous species of related genera of the subtribe
Triphasiinae as well as from the danger of hybridization with vigorous new
genera that evidently arose in rapid succession in southeastern Asia and the
East Indian Archipelago. Thanks to this age-long isolation, even
today the leaflets of Citropsis (and especially the unifoliolate leaves
of certain forms of C. gabunensis) resemble very closely the leaves of
Citrus (and doubtless those of the common ancestor of Citrus
and Citropsis) in shape, texture, venation, and color. That
this resemblance of the leaves is no mere analogy is proved by the fact that
the species of Citropsis, so far as they have been tested, graft readily
on Citrus, and vice versa. Citropsis and
Pleiospermium are the two genera that most nearly represent today the
remote ancestors of Citrus.
The species
of Citropsis are all native to tropical and subtropical Africa, from
Sierra Leone to Angola on the west coast and from Uganda south to Mozambique in
East Africa, as well as to many, if not all, of the regions of central Africa
lying between. As this immense territory includes vast areas still
only inadequately explored, it is highly probable that several, perhaps many,
more species of Citropsis will be discovered in the future.
A striking proof of the inadequacy of our
botanical exploration of tropical Africa is shown by the fact that, of the
eleven species of Citropsis here recognized, eight were discovered after
1900 and five of these after 1920. The last three species to be
found—C. gilletiana, C. tanakae,and C. daweana—were
described as recently as 1940! The two last-named are strikingly
different from all the other known species and from each other.
They are not mere small species made by splitting large species, but are new
and aberrant members of the genus, which when better known may need to have new
subgenera made to include them. One subgenus, Afrocitrus,
has already been named to include three allied species, all growing in western
tropical Africa.
In the Congo, it was found
that Gillet’s cherry-orange, C. gilletiana, the most vigorous species in
the genus, growing to be a tree 10 meters high, could be used to advantage as a
rootstock for cultivated varieties of Citrus because of its resistance to
foot rot (very injurious to cultivated varieties of Citrus
there). This species proved to be an even better rootstock than the
sour orange for the orange, the mandarin, the pummelo, and the lemon (see below). It is one of the more important rootstocks
for Citrus found in this century.
In
view of the beauty of their foliage, the refreshing fragrance of their flowers,
and the attractive aroma of their tiny fruits, coupled with their rapid growth
and free flowering while still small trees, an effort should be made to test
all the species of Citropsis as greenhouse ornamentals in the cooler
climates and as out-of-door ornamentals in subtropical regions. Keys
to the subgenera and species of Citropsis are presented.
Subgenus Citropsis
Subgenus
Eucitropsis Swing. & M. Kell. Jour. Arnold Arbor. 21:125.
1940.
Type.—C.
Schweinfurthii.
Leaflets usually large,
petioles and rachis segments broadly winged; ovary ovoid without a single large
oil gland at the top of each locule; stigma medium-sized, depressed-globose,
without large oil glands; staminal filaments glabrous.
Most of the species of this subgenus have large
leaves, and three of them, C. articulate, C. gilletiana, and C.
latialata, have leaves and leaflets larger than those of any other members
of the tribe Citreae except possibly the palmately trifoliolate leaves of
certain species of Luvunga. The species of this subgenus are
rather sharply distinguished from those of the subgenus Afrocitrus by the
ovary and stigma characters.
1.
Citropsis schweinfurthii (Engl.) Swing. & M. Kell. Jour. Agr. Res.
1:426. 1914. Limonia schweinfurthii Engl. Notitzbl. Bot. Gart.
Berlin 1:28. 1895. Illus. Swingle & M. Kellerman, loc.
cit. figs. 1-2, 4(C)-7, pl. 49; Swingle, in Bailey, Stand. Cycl.
Hort. 2:780, fig. 972. 1914; Engler, Die Nat. Pflanzenfam.
19a:348, fig. 158,K (copy of fig. 4,C above). 1931; fig.
3-24 this work.
Type.—Uando,
northwestern Uganda (Schweinfurthii, No. 3656, 1870). Herb. Bot.
Mus., Berlin-Dahlem.
Distribution.—Northeastern tropical Africa, northwestern Uganda, and
northeastern Republic of the Congo, at altitudes of 600-1,600
m.
Common name.—Uganda
cherry-orange.
Small tree, ultimate branches
at first angled, soon terete, often with single or paired spines 1-2.5 cm long;
leaves medium-sized, 3-5-foliolate, 12-30 X 12-20 cm, leaflets oblong or
lanceolate, tapering rather abruptly to a short, blunt acumen, cuneate at base,
margins denticulate to subentire, terminal leaflets usually narrowly cuneate at
base, 10-15.5 X 3-5.5 cm, lateral leaflets more broadly cuneate at base, 8-14 X
3.5-5.5 cm, petioles broadly winged but usually slowly narrowed and rounded at
both ends, 5.5-10 X 2-3 cm, rachis segment similar in shape to petioles but
shorter and narrower, 4-9 X 1.5-2.8 cm; inflorescences axillary, short (1-2
cm), many flowered, pedicels 2 mm long, 1 mm wide at base, gradually thicker
upward and about 2 mm wide at base of calyx, flowers white, fragrant, usually
4-merous, about 2.2-2.5 cm when fully expanded, calyx with 4 thick, short,
blunt lobes, petals 4, 12-15 X 4-5 mm, broadly rounded at tips, stamens 8,
10-11 mm long, flattened and broad 3/4 of way to top, 0.5 mm thick, 1.4-1.6 mm
broad; disk 1 mm high, 3 mm broad, surrounding narrowed base of ovary with a
shallow (0.3-0.4 mm), thick-walled cup with small scattered oil glands on
inside near base, pistil 10-11 mm long (including disk), ovary ovate,
4-locular, 2 mm high, 2.5 mm wide, rounded at tip with style often (always ?)
attached to one side of center of tip!, style short, stout, 6-7 X 1.1-1.4 mm,
expanded slightly at tip, stigma cushion-shaped, 0.9-1.1 mm high, 2-2.5 mm
wide, 4-lobed at tip, with very small oil glands between stylar canals; fruits
subglobose, lemon-yellow when ripe, fragrant, 2-2.2 cm diam., locules filled
with numerous, sessile pulp-vesicles that occupy all free space, seeds slightly
roughened, 9-12 X 7-8 mm, monoembryonic.
Citropsis schweinfurthii was grown for many years in the United States and
thrived in the Washington greenhouses of the U.S. Department of Agriculture,
where it flowered profusely and set fruit freely while still a small
tree. The leaflets are narrower than those of most of the other
species except the related C. angolensis of Angola and the very
different C. daweana of Mozambique. The leaflets of the
Uganda cherry-orange show 6 to 8 lateral veins, slightly elevated below and
slightly depressed above. The surface of the leaflets is nearly
plane. The fruits of C. schweinfurthii are often borne in
clusters of two to five on short axillary inflorescences; they are almost
spherical or very slightly flattened at the top and 22 to 25 mm in
diameter. The peel is very thin, 1 to 1.5 mm thick. The
ripe fruit ranges in color from Empire yellow to Apricot yellow (Ridgway, 1912,
pl. 4). The pulp, when seen in cross section, is straw yellow,
amber yellow, or even mustard yellow (Ridgway, 1912, pl. 16) near the
peel. The pulp-vesicles fill all the space in the cells of the
ovary not occupied by the plump seeds, of which 1 to 4 may develop, never more
than one in each locule. The pulp-vesicles contain in their center
a long column of oil droplets, irregular in diameter and not sharply
delimited. The oil droplets are extremely variable in size, though
most of them are small. The mawkish taste of the pulp is doubtless
due to these droplets of oil.
The Uganda
cherry-orange can be grafted readily on Citrus (Swingle and M. Kellerman,
1914, p. 435, pl. 49). It can also be made to grow, but less
vigorously, when grafted on the wood apple, Feronia limonia, or on the
Philippine tabog, Swinglea glutinosa. It is well worth
growing as an ornamental in the greenhouse, as it not only has handsome
foliage, but also bears, for months at a time, a great profusion of white bloom
looking much like orange flowers, with a strong but agreeable odor about midway
between that of citrus flowers and that of the North American wild plums
(Prunus americana and other species).
2. Citropsis angolensis Exell, Jour. Bot., Brit. & For.
65(Suppl. 1):53.
Type.—Angola,
Amboine (Grossweiler, No. 4454). Herb. Brit. Mus.
Fragments and serial microtome sections, S. and T. Nos. 273-A, 273-B, and
273-C, 12 slides, in Herb. Natl. Arbor., Washington,
D.C.
Distribution.—Known with certainty
only from the type locality.
Common
name.—Angola cherry-orange.
The original
description by Exell reads as follows: "A tree with brown branchlets; leaves
trifoliolate, leaflets elliptical or obovate, cuneate at base, margins remotely
and irregularly crenate, glabrous on both surfaces, petioles broadly winged,
elliptical; flowers pedicellate, pedicels glabrous, in clusters in the axils of
the leaves; sepals 4, triangular, small; petals 4, oblong, imbricate, rounded
at the tips; stamens 10, filaments free, flattened, glabrous; ovary 4-locular,
style scarcely thickened at the base.
"A tree,
total height 15 feet [4.6 m]; shortly branched from the base.
Leaflets 8-12 X 4-6 cm; petiole 7-8 X 2.5-3 cm; pedicels 5-7 mm long; petals
12-14 X 5-6 mm; filaments of stamens 8-9 mm long; anthers 4 mm long affixed
near the base; ovary 4 mm long and 2.5 mm in diam., style 6 mm long; young
fruits 8 X 5 mm.
"The species is nearest to
C. Schweinfurthii (Engl.) Swing. and M. Kell., but has rather longer
pedicels and a slightly longer and slenderer style—it is, moreover, apparently
devoid of spines."
The two flowers from the
type specimen were kindly donated to Swingle by the Curator of the Herbarium of
the British Museum. A nearly mature flower bud was cut into
several hundred serial microtome sections (both cross and longitudinal) and a
fully mature flower from which the petals and stamens had just fallen was
restored to natural state and photographed. The following
characters were observed:
Pistil slender, 1.2
cm long, with a very slender style, 6 mm long, borne on a slender pedicel, 0.7
mm diam. at base and 1 mm diam. at top; calyx very small (2.3 mm diam.), lobes
triangular, acute, with several (2 or 3) small oil glands near the tip;
stamens free, filaments flattened, anthers 3-3.5 mm long, apparently without
an oil gland near the tip; disk small, 1 mm high, 1.3-1.5 mm wide, and nearly
cylindrical on mature, dry flowers, but bulging to 1.8 mm at mid-height in
nearly mature flower buds (and on a restored mature pistil), not cupulate and
not enclosing the ovary base; ovary truncate-fusiform (ellipsoid), 3.5(-4 ?) mm
high, 1.3 mm wide at base, 1.8 mm wide in middle in dry flowers (2-2.5 mm wide
on a fully restored mature pistil), 4-locular, narrowed at tip and merging
gradually into style base; style slender, 6 mm long, 0.8-1.3 mm wide, expanded
rather abruptly into the subglobose stigma which is 1.1 mm high, 1.6 mm wide
when dry (1.2 mm high, 2 mm wide in restored mature pistil), with 1 or 2 small
oil glands between the stylar canals.
The
flower characters of this species show clearly that, in spite of the general
resemblance of its leaves to those of C. schweinfurthii, it is in fact a
very distinct species, differing decidedly in its spineless twigs, in its
longer ovary and its longer, more slender style arising from the center of the
top of the ovary (not from one side as in C. schweinfurthii), and, even
more strikingly, in its very different, noncupulate disk, which does not
enclose the base of the ovary even to the slightest extent.
This species of Citropsis can, as a matter
of fact, be distinguished only with difficulty from C. schweinfurthii by
any of the easily visible, grosser characters of the leaves, flowers, or fruits
and has in fact been considered by experienced taxonomists to be a mere synonym
of it, not even worthy of being distinguished as a variety. At
first glance the only distinguishing character is the absence of spines in
C. angolensis and their abundance in C. schweinfurthii.
However, when the flowers are studied critically, striking differences
appear which not only separate clearly the Angola cherry-orange from the Uganda
species but also distinguish it from all the other known species of the genus
Citropsis.
3. Citropsis
articulata (Willd.) Swing. & M. Kell. Jour. Agr. Res. 1:433.
1914. Citrus articulata Willd. in Spreng., Syst. Veg. 3:334. 1826;
Limonia preussii Engl. Notitzbl. Bot. Gart. Berlin 1:28. 1895;
Citropsis preussii (Engl.) Swing. & M. Kell. Jour. Agr. Res.
1:423. 1914. Illus. Swingle & M. Kellerman, loc. cit.
p. 424, fig. 3; p. 425, fig. 4,A; Engler, Die Nat. Pflanzenfam.
19a:348, fig. 158,F,G,H.
1931.
Type.—In the mountains 50-75
km north of Accra (Isert. 1786). Herb. Bot. Mus.,
Berlin-Dahlem.
Distribution.—Ghana,
Togo, Cameroon, Republic of Congo (?). This species widely spread
in tropical West Africa, is everywhere a shrub, according to Engler (1931, p.
347).
Common name.—West African
cherry-orange.
A shrub or small tree with
solitary axillary spines, 0.5-2 cm long, even on fruiting branches; leaves
large or very large, 15-33 X 8.5-25 cm, leaflets broadly oval or
oblong-lanceolate, 13-19 X 7.5-10.5 cm, rounded at apex and abruptly narrowed
into a blunt acumen often very short (3-5 mm) but sometimes longer (8-12 mm)
and acute at the very tip, broadly or narrowly cuneate at base, lateral
leaflets slightly smaller than the terminal one, 10.5-15 X 5-7.5 cm, apex like
that of terminal leaflets but base more broadly cuneate or rounded, margins
serrulate or shallowly crenulate, petioles 5.5-8.5 X 1.7-3.5 cm, narrowly
elliptical, usually narrowed at both ends but sometimes broader at the tip or
even obovate (as in the type specimen); inflorescences about 3 cm long (1/3-1/2
as long as the petiole); flowers white, 4-merous, calyx 4 mm wide, lobes
triangular, acute, petals 4, linear-oblong, 18 X 4 mm, stamens 8, filaments 8 X
1 mm, anthers 3.5 mm long, affixed near the base, disk 1 mm high, ovary
4-locular, ovoid-acuminate, 3-4 mm high, 2.5 mm wide, style long (12-15 mm),
slender, expanding gradually and merging with top of ovary, stigma subglobose,
2.5 mm diam., 4-lobed; fruits subglobose with a truncated, conical style base,
1.5-2 mm long, persisting at apex.
This was
the first species of Citropsis to be described. It was
discovered by P. E. Isert, who called it a new species of lemon with jointed
leaves: "une nouvelle espèce de citroniers avec des feuilles
articulées." One of Isert's specimens was sent to the herbarium at
Berlin, and Willdenow named it Citrus articulata in 1826, but did not
describe the flowers, which had perhaps fallen off the "many-flowered
peduncles" he mentioned. In 1895, Engler proposed as a new species
Limonia preussii, found growing on the shores of Elephant Lake in what
is now Cameroon. The description was the first full and accurate
one published of a Citropsis.
The very
defective type specimen of Citropsis articulata, consisting of a short
twig and two petioles in the Berlin-Dahlem herbarium, is all that is known to
be extant of Isert’s material. This type specimen, when studies by
Willdenow, must have been more complete than it is now, for his original
description reads, in translation: "C.[itrus] with large, articulated,
obovate, leaf-like petioles, leaves oblong, peduncles
many-flowered." No inflorescences are now to be seen and no leaves
(or leaflets). Possibly, however, Willdenow mistook detached oblong
petioles for leaves. However, Engler (1915, p. 758), who had access
to the type specimen of Citrus articulata and abundant material of his
own species, Limonia preussii, said: "…indeed I cannot avoid combining my
Limonia preussii [with Citropsis articulata] as the width of the
petiole wing shows no specific differences."
Engler gave C. articulata a very wide range, from the type locality
north of Accra in modern Ghana through near-by Togo (Koli near Kame) east to
what is now Cameroon (Elephant Lake, type locality of Limonia preussii)
and south to the present-day Republic of Congo (Brazzaville) and the Republic of
the Congo (Leopoldville) (including Limonia demeusii De Wild. and L.
poggei Engl., of which the flowers are not yet know). Future
study of flowering material is needed, however, to determine whether C.
articulata has so wide a range or not.
For
the present, Engler's view concerning the identity of C. articulata is
accepted, but flowering specimens from the type locality might even show it to
be identical with C. mirabilis, native to the ivory Coast, adjoining
Ghana to the west, in which event the name C. preussii would be applied
to what is here called C.
articulata.
Citropsis preussii,
here considered as synonymous with C. articulata, is a shrub with very
large leaves, nearly as large as those of C. gilletiana, which grows to
be a tree sometimes 10 meters high.
4. Citropsis mirabilis (A. Chev.) Swing. & M. Kell. Jour.
Agr. Res. 1:432. 1914. Limonia mirabilis A. Chev. Bul. Soc. Bot.
France 58(mem. 8d):144. 1912. Illus. Swingle & M.
Kellerman, loc. cit. p. 425, fig. 4,B (pistil only); Engler, Die
Nat. Pflanzenfam. 19a:348, fig. 158,J (copy of above).
1931.
Type.—West Africa, Ivory Coast,
between Sanroa and Quode on Koué River (an affluent of the Sassandra River)
(Chevalier, 1909, No. 21609). Herb. Chev.,
Paris.
Distribution.—West Africa: Ivory
Coast. Up to now this species was known only from the type
locality.
Common name.—Ivory Coast
cherry-orange.
A much-branched and very spiny
shrub, 2-5 m high, completely glabrous, spines 2-4 cm long; leaves
3-5-foliolate, leaflets oblong or broad-elliptical, 9-16 X 5-9 cm, the terminal
leaflet larger than the lateral ones, apex narrowed into a short, blunt acumen,
base cuneate or badly rounded (when the terminal leaflet is borne singly on a
short rachis segment, as in some 3-foliolate leaves); inflorescences small
(5-10-flowered), axillary racemes, 2-3.5 cm long, pedicels slender, 3-5 mm
long, 0.5 mm wide at base and gradually expanding to 1 mm diam. at top; flowers
white, 4- (rarely 5-) merous, sepals greenish, 1 mm long, petals linear, 13-15
X 4-5 mm, soon falling, stamens free, filaments flattened, disk very short (0.5
mm), forming a loose, very shallow cup (0.2-0.3 mm deep) at base of ovary,
pistil in fully developed flowers 11-13 mm long (including disk), ovary
slender, 2-2.5 X 1-1.2 mm, style 6-9 mm long, slender at base (0.2-0.3 mm
diam.), gradually expanding upward to 0.6-0.7 mm at top, stigma 0.5-0.8 mm
high, 1-1.2 mm wide, 4-lobed, with 2 small oil glands in each space between
stylar canals; fruits (only immature seen) borne on pedicels.
This species much resembles C. articulata,
differing chiefly in the somewhat shorter style not expanding at the base and
not merging into the top of the ovary. Possibly C. mirabilis,
when better understood, will be ranked as a variety of C. articulata
occupying the westernmost part of its wide range.
5. Citropsis gilletiana Swing.
& M. Kell. Jour. Arnold Arbor. 21:116. 1940. Illus. "W. G.
S[mith (?)]," Gard. Chron. 3 ser. 53:378, fig. 159. 1913; Goossens, Bul.
Agr. Congo Belge 15:161, fig. 45. 1924; Swingle & M. Kellerman,
loc. cit. pl. 1, figs. 1-7; pl. 2, figs. 1-5; fig. 3-25 this work.
Type.—Washington, D.C., Bureau of Plant Industry greenhouse, tree grown
from seeds sent by Père J. Gillet from type locality, Kisantu, Belgian Congo
(Swingle, C.P.B. No. 7800 G = P.E.I. No. 109622), flowering branch; also several
paratypes of leafy shoots, flowering and fruiting branches, fruit, etc., cut
from type tree; photographs and serial microtome sections S. and T. Nos. 501-A,
501-B, 501-C, and 501-D, 40 slides with 1,339 sections from type tree, in
Herb. Natl. Arbor., Washington, D.C.
Topotypes.—Belgian Congo, Kisantu, Jardin Botanique (J. Gillet); 2 branches
with very young fruits, March 5, 1913, in Herb. Natl. Arbor., Washington,
D.C.
Distribution.—Reported (Goossens,
1924, p. 162) from Ubangi and Equateur regions of northwestern Belgian Congo
(now Republic of the Congo) and cultivated at the Jardin Botanique
d'Eala.
Common name.—Gillet's
cherry-orange.
Tree 8-10 m tall, young
branches glabrous, 2.5-4 mm diam., internodes 2-6.5 (usually 3.5-4.5) cm long,
spines slender, acute, 2-5 cm long, solitary or in pairs; leaves very large,
3-5-foliolate, 16-37.5 X 10-36 cm, terminal leaflets 9-21 X 5-14 cm, lateral
ones (always smaller than the terminal) 7-17 X 4-9.5 cm, petioles obovate, 6-10
X 3.5-7.2 cm, rachis segments (1 or 2) elliptical or obovate, 3.5-9.5 X 1-5-7.5
cm (usually 3.5-6 X 1.5-4 cm); inflorescences racemose, axillary, short, 5-12
mm long, flower pedicels 3-5 mm long; flowers white, numerous, about 2.5 cm
diam., but often opening incompletely because densely crowded in short
inflorescences 4-parted, calyx lobes triangular, acute, 2 X 2 mm, petals 16-18
X 4.5-5.2 mm, stamens 8, filaments glabrous, flattened, free, disk 0.8-0.9 mm
high, 2.2-2.4 mm wide, cupulate (0.3 mm deep), pistil (including disk) 10-13 mm
long, ovary ovoid or barrel-shaped, 3.5-4 X 2.5-3 mm, 4-locular, locules with 1
ovule, style 4.5-6 X 1.2-1.3 mm, stigma depressed globose, 1.2-1.3 mm high, 2.5
mm wide; fruits subglobose, 24-25 mm diam., with pedicels 7-9 mm long,
lemon-colored when ripe, peel thin (1-1.5 mm), locules with very numerous
pulp-vesicles filled with granules of yellowish wax; seeds white, smooth,
usually 2-3 (often none), ovoid, 9.5-10 X 5-6 mm, testa very hard, hilum ovate,
1-2 mm diam. with smooth edges, monoembryonic.
The type tree of C. gilletiana was grown
in the citrus greenhouse of the former Bureau of Plant Industry from seeds
received from Père Gillet in March, 1913. Unlike C.
schweinfurthii, growing alongside in the greenhouse, the young tree did not
flower for many years and then only sparingly; later it flowered and fruited
profusely. The type tree produced enormous leaves, both
trifoliolate and 5-foliolate. A trifoliolate leaf at the base of a
vigorous water sprout was 28.5 cm long and 31 cm wide. The terminal
leaflet measured 20.5 by 14 cm, the two lateral leaflets 16.5 by 9.5
cm. The largest leaflets have 7 to 10 principal lateral veins,
raised on the under surface and sunken on the upper surface. The
surface of the leaflet is usually more or less bullate because of the upward
curving of areas of the leaf surface limited by the principal lateral veins and
their cross veinlets. The winged petiole, cordate in outline,
measured 8.5 by 7.2 cm. A large 5-foliolate leaf higher up on the
same shoot measured 37.5 cm long and 36 cm wide; the terminal leaflet
(somewhat deformed) was 18 by 10 cm. The two higher lateral
leaflets were 18.5 to 19 by 10.5 to 11 cm. One of the lower
leaflets measured 16 by 10.25 cm (the other was imperfect). The
elongate-elliptical winged rachis segments measured 9.5 by 4.5 cm, the
elliptical winged petiole about 10 cm long and about 7.5 cm wide (one side
defective). These are the largest leaves yet reported on either a
Near-Citrus or a True Citrus Fruit Tree, though they are greatly exceeded in
size by the leaves of some species of Clausena.
In trifoliolate leaves, the terminal leaflet is
usually decidedly longer than the adjoining laterals and frequently much wider,
from one-fourth to one-third or even more; when the terminal leaflet, as often
happens, is borne on a separate winged rachis segment, it is sometimes even
two-fifths wider.
The stigma in cross section
shows eight small oil glands (two between each pair of stylar
canals). The fruits are often crowded, three to six in a dense
cluster in the axils of the leaves of the fruiting branches; they are usually
subglobose, 2 to 2.5 cm in diameter, but are sometimes depressed-globose, 2 cm
long and 2.3 to 2.4 cm wide; they become yellow-orange and are fragrant when
ripe (see fig. 3-25).
Gillet’s
cherry-orange, the largest and most vigorous of all the known forms of the genus
Citropsis, has been reported by Belgian expert horticulturists and
pathologists (Goossens, 1924, pp. 156-62, figs. 42-45: Staner, 1929, pp.
364-66, fig. 170; and Pynaert, 1935, pp. 305-14), on the basis of experiments
carried out at the Eala Botanic Garden near Equatorville in Belgian Congo (now
the Republic of the Congo), to be immune to a very destructive form or foot
rot, supposed to be caused by the brown-rot fungus (Phytophthora
citrophthora), that attacks the lower trunks and upper roots of the species
of Citrus grown in the Congo, including the sour orange, Citrus
aurantium, usually found to be very resistant to foot rot (see Fawcett,
1936, p. 177). It appears that in the Congo foot rot is greatly
aggravated in severity by the larvae of a longicorn beetle, Monohammus
sp. (see Goossens, 1924, p. 159, and Staner, 1929, pp. 365-66), which attacks
first the cambium layer and later the wood of the base of the
trunk. Citropsis gilletiana, besides being immune to the
attacks of the foot-rot fungus, is not attacked by this beetle. It has been found that Gillet's cherry-orange
makes an excellent rootstock for the cultivated varieties of sweet orange,
mandarin, grapefruit, and lemon which were all found to grow more vigorously
when grafted on Gillet's cherry-orange than on sour orange
rootstocks. Furthermore, citrus trees grafted on Gillet's
cherry-orange rootstocks were completely immune to the Monohammus beetle
and also to foot rot, provided the Gillet's cherry-orange stocks were grafted
high enough above the ground to prevent infection from the soil.
Pynaert (1935, p. 313) reported that the native
Citropsis growing about the Eala Botanic Garden in Belgian Congo had been
introduced into Belgium, and that a Jaffa orange grafted on a plant of
Citropsis growing in a greenhouse of the Colonial Garden at Laeken,
Belgium, had made a particularly vigorous growth and in 1935, when still only 2
meters high, had produced a number of fruits. An excellent half-tone
figure of the wild Citropsis found near the Botanic Garden in Belgian
Congo, published by Goossens (1924, p. 161, fig. 45), shows clearly that this
cherry-orange is the same as the one sent to Swingle by Père Gillet from
Kisantu, about 700 kilometers down the Congo River from Eala. It
has the terminal leaflet distinctly larger than the laterals, agreeing with
C. gilletiana (but not with C. latialata). Goossens
reported that near Eala this Citropsis is a tree growing to a height of 8
to 10 meters (26 to 33 feet) that bears tiny, orange-like fruits, 2 to 2.5 cm in
diameter, with a resinous, fragrant pulp in which three or four seeds are
imbedded. The fruits produced by the type tree of Gillet's
cherry-orange growing in the citrus greenhouse at Washington agreed very
closely with those described by Goossens, and the foliage, spines, and fruit
clusters of this type tree resembled exactly what is shown in Goossen’s
photograph of the Citropsis growing wild about Eala.
6. Citropsis latialata (De
Wild.) Swing. & M. Kell. Jour. Wash. Acad. Sci. 28:533. 1938.
Limonia poggei var. latialata De Wild. Ann. Mus. Congo Bot. 5
sér.1:160. 1904. Illus. De Wildeman, loc. cit. pl. 43;
Swingle & M. Kellerman, Jour. Arnold Arbor. 21:pl. 2, figs. 6-10.
1940; fig. 3-26 this work.
Type.—Ikongu village, Sankuru River basin, Lualaba-Kasai district,
south-central Belgian Congo (L. Gentil. No. 1). Herb. Jard. Bot.
l'Etat Brussels; photographs and serial microtome sections, S. and T. No.
397-A, slides 1-6, 429 cross sections, and No. 397-B, slides 1-2, 26
longitudinal sections of a mature flower from the type specimen, in Herb. Natl.
Arbor., Washington, D.C.
Distribution.—Known with certainty only from the type
collection.
Common name.—Ikongu
cherry-orange.
De Wildeman's original French
description of Limonia poggei var. latialata reads, translated, as
follows: "Shrub 3-4 m tall, with slender, cylindric branches, with
imparipinnate leaves, having 2 pairs of lateral leaflets. Leaves
distant 1-5 cm from each other, with the petiole and rachis winged; petioles
3-8.5 cm long and 1.5-5 cm broad, upper [pair of] leaflets distant 2-8.5 cm
from the lower [pair of] leaflets; rachis reaching 4.5 cm in width.
Leaflets oblong, narrowed into a very short petiolule at the base, cuneate at
the apex and at the base, the midrib dividing the leaflet in 2 more or less
unequal parts. Leaf-blades 6-15 cm long and 2-7 cm wide,
irregularly denticulate on the margins; the terminal leaflet regular,
cuneiform at the base, of the same size as the lateral leaflets; axillary
stipules changed into solitary or paired thorns, more or less developed,
reaching 2.5 cm in length. Flowers white, with the odor of orange
[flowers], in axillary or terminal inflorescences; ovaries ovoid, terminated by
a style twice as long, surmounted by a trilobed stigma."
A note states that the type material (L. Gentil,
No. 1) was in flower when collected but "the petals fell off during the
preparation." Evidently the flowers were mature.
A single flower from the type specimen (see
Swingle, 1940c, p. 120) was restored by the modified Juel method,
imbedded in paraffin, and cut into serial microtome sections. First
26 longitudinal sections were cut until a section was obtained exactly through
the center; then the paraffin block was turned at right angles and 429 cross
sections 20 mu thick were cut, including the entire length of the pistil,
disk, calyx, and pedicel. These sections give the following
characters:
Calyx is 4-merous, the lobes about
1.2 mm wide and 1.5 mm long, thick in the middle but with thin edges; disk
shallow, cup-shaped, 0.3-0.4 mm tall and 1.1-1.2 mm wide, pistil 6.4 mm tall,
including the short (1 mm), very shallowly cupulate disk, ovary ovoid, 1.8 mm
tall and 1.6 mm wide, merging rather abruptly into the style, which is 0.8 mm
diam. at the junction with the ovary and nearly 1 mm diam. at the stigmatic
junction; stigma cushion-shaped, 1.5 mm wide and 0.5-0.6 mm high, more or less
4-lobed, with 4 medium-sized oil glands between the 4 stylar canals.
Thanks to the generous loan by Dr. W. Robyns,
director of the National Botanic Garden at Brussels, of one of the very few
flowers of the type specimen, it was possible to clear up the long-standing
confusion of this species with C. gilletiana, from which it can now be
clearly distinguished by many characters of high taxonomic value in this
genus. Citropsis latialata is a shrub only 3 or 4 meters tall
instead of a tree 8 to 10 meters high, it has somewhat narrower leaflets and
the terminal leaflet is of the same size or only very slightly larger than the
adjacent lateral leaflets (in C. gilletiana, on the contrary, the
terminal leaflet is always larger than the lateral ones). The pistil
of C. latialata is only a little more than half as long as that of C.
gilletiana and has a decidedly shorter and much less deeply cupped nectary,
as well as a more flattened stigma with fewer but larger oil glands.
The long-standing confusion of these two species
was due, without doubt, to the very scanty type material of C. latialata
and the lack of any other material of this species from the type
locality. De Wildeman's excellent lithographic plate figuring the
type specimen of C. latialata, natural size, shows five pistils in a
close cluster at the end of a leafy twig. This is fully confirmed
by a photograph of the type filed in the herbarium of the National
Arboretum. These pistils must have been full grown, as the petals
were in place when the material was collected, but fell off during the drying
of the type specimen. As noted above, a single flower from the type
specimen sufficed to distinguish clearly the two species C. latialata
and C. gilletiana.
Subgenus Afrocitrus
Subgenus Afrocitrus Swing. & M. Kell.
Jour. Arnold Arbor. 21:126. 1940.
Leaflets small, acuminate or caudate at apex, petioles and rachis segments
narrowly winged or wingless; ovary clavate or obovoid, with a single very large
oil gland at the top of each locule; stigma subglobose, distended by large oil
glands; staminal filaments sparsely pubescent on inner side.
The three species that constitute this subgenus,
Citropsis gabunensis (type of Afrocitrus), C. zenkeri, and
C. le-testui, are easily distinguished from all the other species of
Citropsis by their long, acuminate leaves, narrowly winged or wingless
petioles and rachis segments, and the striking ovary and stigma
characters.
Citropsis tanakae also has
very small flowers borne on slender pedicels and very slender styles each
ending in a subglobose stigma swollen by several large oil glands, all of
which are characters very like those of the three species named
above. However, C. tanakae differs from them in lacking the
large oil glands at the tip of the ovary, one over each locule, and also in
having simple leaves with very short, wingless petioles, not articulated with
the leaf blade.
7. Citropsis gabunensis (Engl.) Swing. & M. Kell. Jour.
Agr. Res. 1:430. 1914. Limonia gabunensis Engl. Notitzbl. Bot.
Gart. Berlin 1:28. 1895. Illus. Swingle & M. Kellerman,
loc. cit. 425, fig. 4,D-E; De Wildeman, Ann. Mus. Congo Bot. 5
sér. 1:pl. 50. 1904; Engler, Die Pflanzenwelt Afrikas 3:760, fig.
354,L,N. 1915; Engler, Die Nat. Pflanzenfam. 19a:348, fig.
158,L,N (last two, copies of Swingle & Kellerman). 1931; fig.
3-27 this work.
Type.—Sibange Farm,
Menda region near Libreville, Gabon, French Congo (H. Soyaux, No.
105).
Distribution.—Gabon Republic,
Republic of the Congo, and Spanish Guinea.
Common name.—Gabon cherry-orange.
A shrub
or small tree, 1.5-6 m high, ultimate branches slender, 1.5-3 mm diam., with
internodes 2-5 cm long, with single or paired slender spines, 2-2.5 cm long
even on fruiting branches; leaves glabrous, 1-5 foliolate, variable in size,
5-foliolate leaves 15.5-19 X 12-15 cm, 3-foliolate leaves 10-13 X 7-10 cm,
1-foliolate leaves 12-17 X 4-7 cm (including the petiole 2-4 cm long),
leaflets oblong or elliptical, long-acuminate at the apex or caudate, very
variable in size, 1-foliolate leaflets very large, blade 7-14.5 X 3-7 cm,
petiole 2-3.5 X 0.4-1.5 cm, with a broadly rounded base, 5-foliolate leaflets
usually 8-10 X 4-5 cm, but sometimes smaller, 3.5-8 X 1.5-3.5 cm, terminal
leaflet often borne on a short rachis segment and then abruptly narrowed at
base, but if borne at apex of first rachis segment with 2 lateral leaflets then
narrowly cuneate or acuminate at base and longer than lateral leaflets,
petioles variable in size and shape, 1.8-5 X 0.3-2 cm, linear or narrowly
elliptical if narrow, obovate if broad, rachis segments often similar in shape
to petiole, 3.5-5.5 X 0.4-2 cm; inflorescences short, axillary flower clusters,
1.5-2.5 cm long, peduncles apparently unbranched, 8-9 mm long, pedicels long
and very slender, 7-11 X 0.4-0.6 mm (in dry state), short-pubescent, flowers
very small, usually 4-merous, buds 5-6 X 2 mm, open flowers about 10-13 mm
diam., calyx lobes 4, triangular, acute, with a single medium-sized oil gland
near the apex, petals 4, about 6 X 2.5 mm, stamens free, filaments flattened,
sparingly pubescent on inner side; disk very small, about 0.5 mm high, pistil
very short (4-6 mm, including the disk), ovary obovoid, about 1.5 X 0.8-0.9 mm,
3-4-locular with a large, more or less protuberant oil gland at the top of each
locule, style very slender, not expanded at base, about 2 mm long, 0.3-0.4 mm
diam., stigma subglobose, 0.9-1 mm diam., slightly 4-lobed, with about 4 large
oil glands; fruits subglobose, about 1.8-2.2 cm; borne on pedicels 8-12 X 1-2
mm, nearly filled with 3 or 4 ovoid, smooth, hard seeds, 10-12 X 8-10 X 6-8 mm,
with an even-margined hilum 2 X 1 mm, pulp-vesicles almost suppressed the seeds
in most fruits.
This species is the type of
the subgenus Afrocitrus, which includes also C. zenkeri and C.
le-testui. These three species are evidently related to each
other and clearly delimited from the six typical species in the subgenus
Citropsis.
Citropsis gabunensis
has very small flowers, the smallest of any species of the genus. No
other species of Citropsis shows as much variation in the number and size
of the leaflets. The type material from Sibange Farm, Gabon (and
also abundant material collected by Père Klaine near Libreville, Gabon, now in
the Paris herbarium), shows whole branches with very large unifoliolate leaves,
although more often some of the leaves have two or three leaflets.
The typical 5-foliolate form is also common at both places; it often shows a
short winged rachis segment bearing the terminal leaflet, something found only
in trifoliolate leaves of other species of Citropsis. The
large unifoliolate leaves look
astonishingly like those of Citrus in every character—size, shape,
venation, texture, and color; they even have winged petioles very like those of
some forms of Citrus aurantium and C. grandis.
The unifoliolate species of Pleiospermium,
P. sumatranum, P. longisepalum, and P. latialatum, also
have leaves very like those of Citrus and closely resembling those found
on this form of Citropsis gabunensis. Doubtless both
Citropsis and Pleiospermium contain species that resemble closely
remote ancestral forms from which Citrus has evolved.
7a. Citropsis
gabunensis var. lacourtiana (De Wild.) Swing. & M. Kell. Jour.
Arnold Arbor. 21:124. 1940. Limonia lacourtiana De Wild. Ann. Mus.
Congo Bot. 5 sér. 1:159. 1904. Illus. De Wildeman, loc. cit.
pl. 50.
Type.—Bombay, Sankuru
(Lualaba-Kasai), south-central Belgian Congo (L. Gentil, No. 1), Herb. Jard.
Bot. l'Etat, Brussels; photographs and serial microtome sections of type
specimens, S. and T. Nos. 532-A, 533-A, 533-B, 533-C, and 533-D, 18 slides; also
leafy twig and fruit of type collection, in Herb. Natl. Arbor., Washington,
D.C.
Distribution.—Republic of the
Congo: known only from the type locality in the Sankuru River
valley.
Common name.—Sankuru
cherry-orange.
Fruits subglobose, 1.8-2 cm
diam., yellow-orange when ripe, peel 2-2.5 mm thick, rather soft, pulpy tender,
juicy, and of very agreeable flavor, seeds few or none, plump, subglobose,
bluntly conical at one end, 11 X 9-10 mm.
The
type specimen of Limonia lacourtiana was kindly lent to Swingle by Dr. W.
Robyns, director of the Jardin Botanique de l'Etat at Brussels. It
consists of a single fruiting branch collected in May, 1902, with a number of
half-grown and three nearly mature fruits, most of them seedless, but one (or
two?) with a single yellowish-brown (Buckthorn brown, Ridgway, 1912, pl. 15),
rather large, plump, short-ovoid seed, 11 by 8.5 to 9 mm, with a hard testa
having a smooth-edged, ovate hilum 2 by 1 mm. Gentil's original
field label attached to the type sheet calls the plant "un mandarinier sauvage"
and notes its "fruit délicieux." The three to six fruits occur
together in dense axillary clusters and are borne on pedicels 8 to 13 mm long,
which in turn are densely crowded on a very short peduncle only 4 to 10 mm long.
Three topotype specimens in the herbarium of
the National Botanic Garden at Brussels collected nearly a year later by Em.
and M. Laurent consist of two sterile branches with paired axillary spines, 2
to 2.5 cm long, and a fruiting branch from a small tree, 6 to 7 m high, looking
extremely similar to the type specimen, with eight immature fruits, all but one
of them seedless.
Flowers of the variety are
unknown, but a specimen in the Brussels herbarium from the same general Sankuru
region, collected by A. Sapin in September, 1906, consists of two flowering
twigs with axillary clusters containing three to six very small flower buds,
about 5 by 3 mm, with slender pedicels, 0.4 to 0.6 mm diam. These
have pistils (immature?) which when restored by the modified Juel technique
measured 4.4 mm long (including the disk); disk 0.6 to 0.7 mm high, 1 to 1.25
mm wide, cupulate, enclosing narrowed base of ovary for 0.2 to 0.25 mm; walls of
the cup unusually thick (0.4 to 0.6 mm), containing six to ten medium-sized oil
glands arranged approximately in a ring at a level even with or slightly below
the bottom of the cup; ovary 3-locular, 1.3 mm long, 1.1 mm wide at top, 0.5 mm
wide at bottom, with a medium-sized oil gland at the top of each locule; style
2 mm long, 0.5 to 0.6 mm wide; stigma 1 mm long, 1.2 mm wide, with large oil
glands between the stylar canals. The anthers have a single large
oil gland near the tip of the connective. These measurements and
the disposition of the oil glands in the ovary, stigma, and anthers agree very
closely with those found in the flower buds of a specimen of typical C.
gabunensis preserved in the herbarium of the Museum of Natural History at
Paris, collected by Père Klaine at Libreville, Gabon. However,
Sapin’s material from Sankuru has glabrous staminal filaments, whereas those of
the species have scattered, slender hairs on the side facing the ovary, as do
those of C. le-testui. If this absence of pubescence on the
filaments of the variety is found to be a constant character, it will be useful
in separating it from the species at seasons when no mature fruits are to be
found.
The two species of Citropsis
that have and fruited in the United States, C. schweinfurthii and
C. gilletiana, both produce an abundance of small orange-like fruits, but
they are not edible, as the pulp-vesicles contain numerous granules of a waxy
substance of disagreeable flavor. Doubtless C. gabunensis,
belonging as it does to a very different group (the type of the subgenus Afrocitrus discussed above), does not produce this
ill-flavored wax. The species itself has fruits so full of plump
seeds as to leave very little space for pulp. The variety
lacourtiana, on the contrary, has seedless fruits that are filled with
high-flavored pulp.
8.
Citropsis zenkeri Engler, Die Pflanzenwelt Afrikas 3(Heft 1):760.
1915. Illus. Engler, loc. cit. p. 759, fig. 354,A-E;
Engler, Die Nat. Pflanzenfam. 19a:348, fig. 158;A-F. 1931.
Type.—Bipindi, southern Cameroons
(Lat. 3° 20' N., Long. 11° 45' E.). Herb. Bot. Mus.,
Dahlem-Berlin.
Distribution.—Known only
from the type locality.
Common
name.—Zenker's cherry-orange.
Fruiting
twigs with slender, acute, solitary, axillary spines, 1.5-2.5 cm long and 2-2.5
mm diam. at base; leaves 5-foliolate, 25 X 23 cm, leaflets 11-13 X 5-5.8 cm,
ovate-acuminate or short-caudate, acumen 15-20 X 4-1 mm, acute at apex, base
broadly cuneate, with very short (1-2.5 mm) petiolules, petiole 5.5-6 cm long,
narrowly winged, 3-4 mm wide on either side of the midrib, rachis segments
about 5 cm long, narrowly winged, total width at broadest place, 8-9 mm;
inflorescences short, axillary clusters of numerous flowers with pedicels 3-4 X
0.5-0.7 mm, borne in dense clusters (5-8?) on short, unbranched, glabrous
peduncles, 3-4 mm long; flowers medium-sized, 15-18 mm broad when open,
4-merous, calyx lobes deltoid, bluntly pointed, roughened on the back by
numerous oil glands, petals tapering regularly to an acute apex, 8-10 mm long,
with numerous oil glands, stamens 5-6 mm long, anthers 3-3.5 mm long, pistil
7.5-8 mm long (including short, narrow disk), ovary obovoid, 4-locular, 2-3 mm
long, 0.6-0.8 mm wide at base but expanded at apex to 1.5-2 mm in width by 4
large protuberant oil glands (1 at the top of each locate), style cylindrical,
not expanded at junction with the tip of the ovule, 3.5-4 mm long (including
the stigma), 0.4-0.5 mm wide, stigma subglobose, 1.5 mm diam., obscurely
4-lobed, swollen with 4 large oil glands.
This
description is based partly on the short original description but mostly on the
excellent illustrations that accompany it (Engler, 1915), which are made more
valuable by comparable figures of five other species of Citropsis.
The Zenker's cherry-orange is evidently closely
related to C. gabunensis and to C. le-testui, but has even larger,
more protuberant oil glands at the top of the ovary above each
locule. The fruits are unknown, but should be collected in the hope
of finding an edible form with few seeds like C. gabunensis var.
lacourtiana.
9.
Citropsis le-testui Pellegrin, Bul. Mus. Hist. Nat. Paris 27:446.
1921. Illus. Pellegrin, Mém. Soc. Linn. Normandie 26:42, fig. 3.
1924.
Type.—Ndougou in Ngouyé Valley,
French Congo (Lat. 2°-3° S., Long. 10° 30' E.) (Le Testu, No.
2286). Herb. Mus. Hist. Nat., Paris; photographs of type, also 2
leaflets and 3 flower buds of type material with serial microtome sections of 1
bud, S. and T. Nos. 708-A (9 slides, cross sections), 708-B (3 slides,
longitudinal sections), in Herb. Natl. Arbor., Washington,
D.C.
Distribution.—Known only from the
type locality.
Common name.—Le Testu's
cherry-orange.
The original description,
translated (and slightly amended from the equally detailed French redescription
published (3 years later, in 1924) reads: "A small tree, 3 meters high,
branches gray-green, glabrous, slender, more or less spiny. Spines
1 or 2, erect, axillary, 2-3 cm long. Leaves usually 3-foliolate,
sometimes 5-foliolate ([the two pairs] separated 3-6 cm). Petiole
nearly terete, glabrous, 6-8 cm long (or in 5-foliolate leaves, 12-15 cm long),
articulated with the rachis segment. Leaflets ovate, 6-12 cm long,
3.5-7.5 cm wide, apex attenuate-acuminate, acumen obtuse, 1-2 cm long, abruptly
contracted at base into a petiolule 3-4 mm long, glabrous, rigid, membranaceous,
glandulose-punctate, midrib prominent below, depressed above, 6 primary
lateral veins on each side, ascending and anastomosing far from the margin
then curving to meet it, the smaller veinlets reticulate, slender, prominent
below in dried specimens. Inflorescences short, axillary panicles,
very shortly tomentose, peduncles 0.8 mm long, pedicels 5 mm long.
Bracts ovate, acute, 1 mm long. Calyx thick, glandulose,
shortly-villose without, 4-lobed, lobes deltoid, acute, 2 mm long with scarious
margins. Petals 4, glandulose, glabrous, oblong, concave, apex
subangulate, 7 mm long, 3.5 mm wide. Stamens 8, filaments
linear-lanceolate, flattened, shortly-villose, 4 mm long, anthers introrse,
glabrescent, 4 mm long, oblong-saggitate [sic], apex acute and
long-acuminate. Disk prismatic, apex somewhat 4-lobed, 1 mm high,
glandulose, glabrous. Ovary glandulose, glabrous, obpyramidal,
somewhat 4-lobed at the apex, 2 mm high, 4-locular, locules 1-ovulate; style
terete, glabrous, 2-3 mm high, stigma capitate, somewhat 4-lobed.
Fruit subglobose, slightly flattened at the apex, 1.5 cm high, 2 cm broad."
Pellegrin in his comments following the
description stated that the flowers are white, the leaves usually
3-foliolate. Citropsis le-testui is a small tree, 3 m high,
relatively abundant on the slopes of the hills, and never seen growing in wet
situations; the fruit is yellow when ripe.
This remarkable species differs from all others of the genus Citropsis in
having completely wingless petioles and rachis segments, which gives the leaves
a curious aspect. As noted by Pellegrin, the leaves of this species
much resemble those of Balsamocitrus dawei, a very different plant of the
subtribe Balsamocitrinae, not at all closely related to Citropsis.
The figure of C. le-testui cited above
shows the stamen to be 8 mm long, the filament flattened, about 4 mm long, 0.75
mm wide, 0.5 mm thick at the widest part, and short-pubescent on the inner
side. The anther is shown to be about 3 to 3.2 mm long, 1 mm wide,
and 0.75 mm thick, tapering to a sharp point at the apex.
Fortunately, a flower bud ready to open was
available for study from the type specimen. After being restored by
the modified Juel technique and cut into serial microtome sections, the
following characters of the flower parts were observed:
Nearly mature flower buds 9-10 X 4-5 mm; calyx
lobes 3 (4?), 1 mm wide, 1.5 mm long, subacute, much thickened by numerous oil
glands which are larger and more closely juxtaposed in the free lobes; corolla
3- (4-?) merous, petals linear, 9-10 X 5 mm, with sparsely scattered,
medium-sized oil glands; stamens 8, about 7 mm long, filaments free, 4-4.5 mm
long, subclavate, about 1 mm wide, thickest near the apex (0.4-0.5 mm), with
many slender hyaline, scattered hairs on the ventral surface (facing the
ovary), anthers 3-3.5 mm long, 0.6-0.8 mm wide, with a large, elliptical,
acute-pointed oil gland at the apex, 1 mm long, 0.3-0.4 mm wide; pistil (in
nearly mature bud) 7 mm long (including the disk), disk 1-1.2 mm long, cupulate
and enclosing base of the ovary for about 0.4 mm, without noticeable oil
glands; ovary 3- (4-?) locular, obconical, 2.6-3 mm long, 0.5-0.6 mm wide below,
at junction with disk (at bottom of cup), tapering regularly upward, 2 mm wide
at flattened top, which has 1 large, ovoid, somewhat protuberant oil gland
(0.6-0.8 mm diam.) over each locule and numerous smaller oil glands between
them; style arising abruptly, narrowed and slightly countersunk in the center
of the flattish upper face of the ovary, 3-3.2 mm long, 0.5 wide at base,
gradually expanding to 0.8 mm wide at junction with the stigma, sparsely
covered with small, scattered, superficial oil glands; stigma subglobose, 1.2
mm high, 1.5-1.6 mm wide, with several (4-5?) large oil glands, 0.9-1 mm
vertical diam. and 0.3-0.6 mm transverse diam., sometimes smaller (through
pairing).
Thanks to this authentic type
material, it is possible to assert definitely that the pistil of C. le-testui
is very like those of C. gabunensis and C. zenkeri, and in
general is intermediate between them in its characters. The
flat-sided ovary and disk described and figured by Pellegrin (l.c.) were
doubtless distorted by shrinking suffered during desiccation.
SPECIES INADEQUATELY KNOWN AND OF UNCERTAIN RELATIONSHIPS
10. Citropsis tanakae Swing. & M. Kell. Jour. Arnold
Arbor. 21:121. 1940. Illus. Tanaka, Kankitsu No Kenkyû (Citrus
Studies) 78, fig. 71, left (as Citropsis citrifolia, a nomen
nudum). 1933; Swingle & M. Kellerman, loc. cit. pl. 3, figs. 1-5.
Type.—Africa, Sierra Leone
(Afzelius). Herb. Univ. Uppsala, Sweden; photographs and serial
microtome sections, S. and T. No. 226-A, 3 slides, in Herb. Natl. Arbor.,
Washington, D.C.
Distribution.—Known
only from the type locality.
Common
name.—Sierra Leone cherry-orange.
Ultimate
branchlets slender, 1.5-3 mm diam., with solitary, axillary, slender, short
spines, 2-10 mm long; internodes 2-3 (or 4?) cm long; leaves simple! broadly
lanceolate, tapering gradually, or slightly acuminate, into a short, thick,
blunt acumen, 8-10 X 6-3 (or 4) mm, base broadly cuneate, with the margins
slightly decurrent into upper part of petiole, lateral veins numerous, 10-12
on each side, arising at angles of about 75°-80° with the midrib, margins
regularly but shallowly crenulate on upper half, subentire below; petioles very
short (3.5-4 mm), wingless, slender (1-1.5 mm wide), glabrous, not articulated
with the leaf blade; inflorescences very short, few-flowered, axillary; flower
buds small (immature?), about 8 X 3 mm, borne on short, slender pedicels, 2-3 mm
(or more?) long, 0.5-0.75 mm diam.; calyx roughened with numerous oil glands,
glabrous, calyx lobes 4, short (0-3-0.4 mm), with thin scarious edges, with a
single rather large oil gland near the subacute tip, petals 4, about 7 X 3 mm,
glabrous, with many medium-sized oil glands, more abundant at the pointed tips,
stamens 8, about 5.5-6 mm long, staminal filaments glabrous, somewhat flattened
radially, broad at the base where they cohere in groups of 2 or 3 for some
distance (about 2.8 mm), anthers about 3 mm long with a single small oil gland
near the top of the connective; disk cylindrical, glabrous, short, shallow
cup-shaped, fitting rather closely over the base of the ovary for about 0.1 mm,
with a few small oil glands in walls of cup, pistil 5.6 mm long (including
disk), ovary 4-locular, ovoid, 0.93-1 mm long, 0.8-1 mm wide, rounded at top,
without a large oil gland over each locule, style long and slender (4.6 mm long,
0.3-0.4 mm wide), contracted where it joins the ovary in which it is very
slightly countersunk, slightly expanded at the apex where it merges into
depressed globose stigma, 0.7 mm high and 0.9 mm wide, containing several (5-7)
large oil glands.
This species is known only
from a single twig, about 20 cm long, with seven leaves and a single flower
bud, collected in Sierra Leone between 1794 and 1796 by the Swedish botanist
Afzelius, who identified it as Citrus medica, doubtless because the
petioles were not articulated with the leaf.
This unique flower bud and a single leaf were kindly sent to Washington by the
custodian of the herbarium of the Botanical Museum of the University of
Uppsala, Sweden. The flower bud was cut into 454 serial cross
sections, each 20 mu thick, by a modification of the method published by
Juel (1918, p. 14) while he was professor of botany at the University of
Uppsala. This very old material could not be restored as well as
usual, but nevertheless it was possible to work out the flower structure in
minute detail (see Swingle, 1940c, pl. 3, figs. 1-5) and to draw up the
preceding description, which proves beyond doubt that this plant is not a
species of Citrus but a new species of Citropsis.
This remarkable species is unique in the genus
because of its simple leaves with very short wingless petioles (only
one-twentieth as long as the leaf blade and not articulated with
it). All other species of Citropsis have odd-pinnate leaves,
trifoliolate to 7-foliolate or occasionally (in C. gabunensis)
unifoliolate leaves, with petioles that are, however, very evidently winged and
never less than one-eighth as long as the leaf blade. In C.
le-testui the petioles (and rachis segments) are wingless, but are very many
times longer.
The leaves of C. tanakae
are very like the leaflets of the other species of Citropsis, which
always have very short, wingless, nonarticulated petiolules.
Citropsis tanakae has flowers somewhat resembling those of the species in
the subgenus Afrocitrus in having a subglobose stigma distended with large oil
glands borne on a very slender style, but the ovary lacks the large oil glands,
one at the top of each locule, such as characterize the species of this
subgenus.
Besides being one of the most
distinct species of the genus, C. tanakae is also the northernmost
species in Africa, occurring some 500 to 600 kilometers northwest of C.
mirabilis, a very different species, native to the Ivory Coast in West
Africa.
This beautiful Sierra Leone
cherry-orange has been named in honor of Professor Tyôzaburô Tanaka, who, as
soon as he saw the type specimen in the Uppsala herbarium, recognized that it
belongs, not to the genus Citrus, as Afzelius supposed, but to
Citropsis.
11. Citropsis daweana Swing. & M. Kell. Jour. Arnold
Arbor. 21:123. 1940. Illus. Tanaka, Kankitsu No Kenkyû (Citrus
Studies) 77, fig. 70, right (sub nomen nudum, Hesperethusa villosa).
1933; Swingle & M. Kellerman, loc. cit. pl. 3, fig. 6; fig. 3-28 this work.
Type.—Madanda Forest, Portuguese East Africa (Dawe, No. 443).
Herb. Brit. Mus., London; photograph and fragments in Herb. Natl. Arbor.,
Washington, D.C.
Distribution.—Known
only from the type locality (now
Mozambique).
Common name.—Mozambique
cherry-orange.
A shrub or small tree, 3 -5 m
high, branchlets at first angular, soon cylindrical, 2 -4 mm diam; internodes
2-3 cm long; spines axillary, solitary, short, straight, 1-2.5 cm long, 2-4 mm
diam. at base, shorter ones blunt-pointed, the longer ones (1.5-2.5 cm)
sharp-pointed; leaves 5-7-foliolate; leaflets elliptical to rhomboid or ovate,
bluntly rounded at the apex, terminal one narrowly cuneate at base, 5-5.8 X
2-2.4 cm, lateral ones broadly cuneate or broadly rounded at base, 2-4.5 X
1.5-2.4, with 4-6 rather inconspicuous veins, sparsely and evenly dotted with
small translucent oil glands, sparingly pubescent on both sides and on the
margins with very slender, colorless ciliate hairs, margins finely and
shallowly crenate or subentire, petiolules very short (1-1.8 mm); petioles
nearly or quite wingless. 1.2-2.2 cm long, 1-1.5 mm wide; first
rachis segments 1.5-2 cm long, with spathulate wings, 3-6 mm wide and rounded
at the distal end, usually narrowed to a subacuminate base at the proximal
end, second rachis segments elliptical, broader (about 2.5 X 0.8 cm), not
spathulate; flowers and fruits unknown.
This
remarkable species is known only from the type material collected by M. T. Dawe
(when director of agriculture of the Companhia de Moçambique) in the Madanda
Forest, a rubber-producing region, which lies 100 to 200 kilometers to the
southwest of Beira, between the Lucite River (Lat. 20° S.) and the Save River
(Lat. 21° 30' S.), to the west of the Sofala lands (about Long. 34° E.) and to
the east of the Mossurize district (about Long. 33° E.).
Unfortunately the flowers and fruits of this
species are unknown; however, the leaf characters are so distinctive that it
cannot be mistaken for any other Citropsis. The leaves,
including the petioles and rachis segments, are sparingly pubescent on both
surfaces, whereas in all the other species they are glabrous; the leaflets are
the smallest known in the genus. The petiole-rachis wings become
progressively wider toward the tip of the leaf, that is, the petiole is
wingless or nearly so, the first rachis segment narrowly winged (wider at the
tip), and the second segment still wider. This is just the reverse
of what is found in the other winged species of Citropsis.
The scanty type material seems to show Kurztrieben, like the leaf and
flower spurs of Poncirus trifoliata and some of the Hard-Shelled Citroid
Fruit Trees (subtribe Balsamocitrinae).
The
leaf, petiole, and rachis characters of this species show a great similarity to
those of certain forms of Hesperethusa crenulata from peninsular
India. However, as noted above, certain forms of Citropsis gabunensis have leaves
and petioles greatly resembling those of Citrus but have flowers and
fruits unmistakably belonging to Citropsis. Only the study of
flowers and fruits can reveal the exact relationships of C. daweana.
The Mozambique cherry-orange has the appearance
of being a somewhat xerophytic plant; the other species are evidently
mesophytes and usually grow in tropical rain forests.
This is without question the least known species
of the genus Citropsis and at the same time the most distinct
one! When flowers and fruits are available for study it will very
probably be found to constitute a new subgenus, if not a new genus.
It is of great phylogenetic interest as a measure of the nature and rate of
evolution that has occurred since Citropsis was cut off from any
connection with its relatives in southeastern Asia, eastern India, and the
Monsoon region.
XX. Atalantia
Corrêa
XX. Atalantia
Corrêa, Ann. Mus. Hist. Nat. Paris 6:383. 1805. Rissoa Arn. Nov.
Act Acad. Caes. Leop.-Carol 18:324. 1836; Malnarega Raf. Sylv.
Tell. Mant. 143. 1838.
Type
species.—Limonia monophylla Roxb. non (L.) = Atalantia
monophylla DC.
Distribution.—Southwestern Asia: India, Ceylon, Burma, Cambodia, Laos,
North Vietnam, South Vietnam, southern China; also Sumatra, Banka (?), Java
(?).
Common name.—Atalantia.
Small trees or shrubs, twigs more or less angled
when young, soon cylindrical, glabrous in some species, more or less hairy when
young, with single, stout, sharp spines in the axils of the leaves, or
spineless, especially fruiting branches of old trees (some species [A.
rotundifolia] nearly spineless); leaves 1-foliolate (or rarely simple),
more or less coriaceous, strongly veined, and usually with conspicuous
reticulations between the lateral veins; petioles wingless, often pubescent
when young, articulated with the leaf blade, short, often less than 1/10 as
long as the leaf blade; flowers small, axillary or rarely terminal, fascicled,
or in racemose corymbs or panicles; calyx 3-5-lobed, or forming a continuous
envelope over the corolla, stamens, and pistil, and splitting more or less
irregularly into 2 or 3 parts as the flower expands; petals imbricate; stamens
6-10, free, or more or less connate into a staminal tube which is sometimes a
closed cylinder with the anthers attached to the rim; anthers short, broadly
ovoid; disk annular, short; ovary ovate, or subglobose, with 3-5 locules, with
1 or 2 ovules in each locule; fruits small, subglobose, with numerous
broad-based, sessile, slender, tapering pulp-vesicles filling all the space in
the segments left by the seeds; peel thin, greenish-yellow on ripe fruit,
dotted with oil glands; seeds oblong, green within, sometime polyembryonic.
The genus Atalantia contains small trees
somewhat resembling Citrus in general aspect, bearing fragrant white
flowers and globose fruits with the appearance of diminutive greenish-yellow
oranges. The pulp-vesicles are, however, different from those of
Citrus in being sessile instead of stalked. The leaves of
Atalantia, although almost always unifoliolate like those of
Citrus, are very different in having much more prominent, more numerous
lateral veins, and veinlets forming reticulations between the lateral veins.
Besides the typical species of Atalantia,
which have numerous sessile, conical pulp-vesicles, there are at least two
species which belong to the subgenus Rissoa, based on the type species
A. ceylanica. This species has very large seeds which almost
completely fill the locules of the fruit, leaving very little space for
pulp-vesicles. Atalantia rotundifolia is closely related to
A. ceylanica and is perhaps a spineless brachytic mutation of
it. A third species, A. guillaumini, somewhat resembles A.
ceylanica and like it has large seeds almost completely filling the locules
of the fruit. However, so far as could be determined from the
scanty fruits examined, no pulp-vesicles are present. It may be
found that there has been an evolutionary tendency of the species of the
subgenus Rissoa to lose the pulp-vesicles, resulting in the development
of completely pulpless species obviously closely related to Atalantia in
spite of lacking one of the most important characters of the genus and even of
the subtribe Citrinae.
It has in the past been
supposed that the genus Atalantia was founded by Corrêa da Serra to
include Limonia monophylla L., now believed to be a synonym of A.
ceylanica. Corrêa cited no species under his new genus
Atalantia but almost certainly had in mind the "Limonia monophylla
L." described and figured by Roxburgh (1795, vol. 1, pp.59-60, pl. 83), since he
spoke in detail of "singular monadelphy" of the stamens with the filaments
fused into a complete cylinder, just as had been figured by Roxburgh ten years
previously. That he was familiar with Roxburgh's great work on
Plants of the Coast of Coromandel is shown by the fact that immediately
after discussing the genus Atalantia he mentioned Limonia arborea
and L. pentaphylla of Roxburgh (two species on which Corrêa based his
new genus Glycosmis)and cited Roxburgh's plates of them which follow
immediately after the plate of Limonia monophylla.
In view of these facts, it appears that the genus
Atalantia is, in fact, based on Roxburgh's description and illustration
of a plant which he thought was Limonia monophylla L., but which is now
known to be another species, very probably Atalantia monophylla
DC. Keys to the subgenera and species of Atalantia are
presented.
Subgenus Atalantia
Subgenus
Euatalantia Swing. in Bailey, Stand. Cycl. Hort. 1:426.
1914.
Type.—Atalantia monophylla
DC.
Stamens connate, forming a cylindric
tube, or connate in groups, rarely free; fruits with numerous well-developed,
juicy pulp-vesicles that fill all the space in the fruit not occupied by the few
medium-sized seeds; stipuloid paraphylls absent.
Seven of the species of Atalantia fall in
this subgenus. In general they much resemble the species of the
subgenus Rissoa.
1.
Atalantia monophylla DC. Prodr. 1:535. 1824. Limonia
monophylla Roxb. (non L.) Pl. Corom. 1:59. 1795; (?) Turraea
virens Hellen. (non L.) Act. Holm. 9:308. 1788; (?) T. spinosa
Willd., in L., Sp. Pl. ed. 4/5. 2:554. 1899 (= T. virens
Hellen.); Atalantia spinosa (Willd.) Tan. Jour. Bot. Brit. & For.
68:232. 1930 (name preoccupied!); (?) Mal-naregam, Rheede, Hort.
Malab. 4:27. 1679; (?) Malnarega malabarica Raf. Sylv. Tell. Mant.
143. (1838); (?) Atalantia malabarica (Raf.) Tan. Jour. Indian Bot. Soc.
16:233. 1937; Atalantia floribunda Wt. Icon. Pl. Ind. Or.
4:16. 1850. Illus. (?) Rheede, loc. cit. pl. 12; (?)
Hellenius, loc. cit. pl. 1, fig. 1; Wight, loc. cit. pl. 1611.
1874; Beddome, Fl. Sylv. Anal. Gen. pl. 7, fig. 5. 1871; Engler, Die Nat.
Pflanzenfam. 3(pt. 4):fig. 111,C-D. 1896; ibid.
19a:fig. 150,C-D. 1931; fig. 3-29 this work.
Type.—British India, Coromandel (Roxburgh). Brit.
Mus.?
Distribution.—Common in India
(except in Himalayan region and in Bombay State), Ceylon, Burma, Thailand,
Cambodia, Laos, North Vietnam, South Vietnam.
Common name.—Indian atalantia.
A small, much-branched tree with rounded twigs
usually slightly hairy at first, becoming glabrous, with single, stout, sharp
spines, 10-15 or 20 mm long, or unarmed; leaves bright green above, paler and
reticulate-veined below, ovate-lanceolate or elliptical, variable in size, 3-15
X 2-4 cm (usually 4-8 or 10 X 2.5-3.5 cm), margins entire or slightly undulate,
apices obtusely rounded, often emarginate, bases broadly cuneate, articulated
with the wingless petioles that are 5-10 mm long and 1.5-2 mm wide, flattened
above, veins strongly marked on the under surface, 10-14 on each side, branched,
forming reticulate veinlets and also forked near the margins and anastomosing;
flowers long-pediceled on short racemes (or clustered) in the axils of the
leaves, pedicels 6-15 mm long, finely pubescent or glabrous, merging into the
calyx; calyx more or less irregularly and deeply 2-cleft, and glabrous or
finely pubescent; corolla 4-5-merous, petals white, glabrous, bluntly rounded
at apex, 8-10 mm long; stamens 8-10, united into a tube 6-8 mm long with the
anthers borne on the free tips of the filaments; pistil slender, 6-7 X 1 mm,
ending in an abruptly expanded, 3-4-lobed stigma, 1.2-1.5 mm diam.; style
caducous; fruits globose, 1.5-2 cm diam., yellowish-green when ripe, locules
filled with numerous cuneate, sessile pulp-vesicles which are broad-based and
tapering to a point at their free end, toward the center of fruit; seeds
usually only 1 to a locule; often some of the locules do not develop seeds.
The most remarkable character of this species
(found also in the closely related species A. macrophylla) is its very
curious calyx. This is a hollow, balloon-like envelope, not
articulated with the pedicel and without any sign of sepals. Soon
this undifferentiated envelope splits more or less irregularly into 2 (rarely
3) lobes, and exposes the corolla, stamens, and pistil to view.
The nomenclature of this species is in a confused
state. Limonia monophylla of Linnaeus has been assumed by
almost all taxonomists to be this species. However, Trimen (1893, p.
227) listed L. monophylla L. as a synonym of Atalantia ceylanica,
and stated that "the figure of Burmann [1737, pl. 65, fig. l]…appears to
represent this rather than A. monophylla…Hermann's specimen…is certainly
A. zeylanica [ceylanica]." Airy-Shaw (1939, p. 291)
again examined Hermann’s specimen (which is in fact the type specimen of
Limonia monophylla L.) in the British Museum and saw no reason to doubt
Trimen's findings. Airy-Shaw then showed that A. monophylla
was based by De Candolle expressly on "Limonia monophylla Linn." of
Roxburgh (1795, vol. 1, pp. 59-60, pl. 83) and that L. monophylla L. was
not cited. However, the plant described and figured by Roxburgh was
certainly not A. ceylanica and hence was not Limonia monophylla
L. Airy-Shaw assumed that Roxburgh's plant from the Coromandel Coast
was, in fact, A. monophylla as understood by Hooker (1875, p. 511), and
accepted the name A. monophylla DC. for it. However, Roxburgh
described and figured a plant having a "four-five parted" calyx, a character not
found in A. monophylla as understood by Hooker. That species
has, instead of a normal calyx, a bladder-like continuous membrane covering the
flower bud which, as the bud expands, splits irregularly. Possibly
both Roxburgh and the artist who made the beautiful colored plate of the
Coromandel plant failed to see the irregularly ruptured calyx and, without
careful study, assumed that it was 4- or 5-parted as in most of the other
species of Atalantia and other related genera. Roxburgh
described the leaves as "end-nicked" and "from two to three inches long, and
one or one and a half broad"; nevertheless he published a plate showing a
flowering branch with 23 leaves, none of them emarginate at the apex, measuring
1 7/8 to 2 9/16 inches long, and 5/8 to 1 1/16 inches wide, only one leaf
slightly exceeding the minimum width given in the description and more than
three-fourths of them narrower than 1 inch! In view of these facts,
no great importance should be attached to the fact that neither Roxburgh nor
the artist noticed at that time the abnormal splitting of the calyx, something
no botanist had noticed in any species of Atalantia or in any other plant
belonging to the orange subfamily.
For the
present, it seems best to assume that Roxburgh was describing and figuring the
true A. monophylla of Hooker and that A. monophylla DC. is the
oldest valid name for it.
Atalantia
monophylla has very hard wood that has been recommended as a substitute for
boxwood (Gamble, 1915). The fruits are said to yield a warm oil
used in treating chronic rheumatism by native doctors. One Indian
author (Nadkani) stated that the fruits "make a nice pickle" (Lewis, 1934, p.
80).
When watered frequently, this species
makes a luxuriant growth in the greenhouse (see fig. 3-29). It should be tested as a hedge plant in
warm subtropical climates and in dooryard gardens as an ornamental, since it
has dense deep-green evergreen foliage and bears a great profusion of fragrant
flowers and many small globose yellow fruits. Atalantia
monophylla can be grafted on Citrus, and vice versa.
2. Atalantia macrophylla
(Oliv.) Kurz. Jour. Asiat. Soc. Bengal 44(2):136. 1875; idem, For.
Fl. Burma 1:195. 1877. Atalantia monophylla var.
macrophylla Oliv. Jour. Linn. Soc. Bot. 5(Suppl. 2):24.
1861.
Type.—Tenasserim
(Helfer). Herb. Bot. Gard., Calcutta
(?).
Distribution.—Southern Burma:
Tenasserim; Andaman Islands; also Bangka Island and Java (fide
Engler).
Common name.—Giant Andaman
atalantia.
This species was at first
considered to be a subspecies of A. monophylla, since it has, like that
species, the calyx irregularly split, usually to the base on one side; it
differs, however, in having 2 ovules in each locule of the ovary (A.
monophylla usually has only 1), and in having larger and broader leaves
(5-15 X 3.5-6.5 cm). Kurz (1877, vol. 1, p. 195), who raised A.
macrophylla to specific rank, found that it had fruits much larger than
those of A. monophylla; he stated that they are "the size of a wood
apple" [which measures about 3.5-5 cm diam.]. The pedicels of the
short racemes are long (6-15 mm), as in A. monophylla, but are very
stout, stiff, and compressed. Atalantia macrophylla is a
small tree, 8-10 m in height, occurring in the Andaman Islands and
Tenasserim. Kurz described it as glabrous in all parts and almost
spineless.
Unfortunately, this very
interesting Atalantia has been very inadequately studied and has never
been figured. Kurz showed clearly that the two species agree in
possessing this remarkable character by putting both A. monophylla and
A. macrophylla into the first division of his key to the species of
Atalantia, which reads: "Calyx irregularly lobed, split to the base on
one side." All other Aurantioideae have flower buds with a more or
less regularly (usually 3 to 5-) lobed calyx. This character,
shared in common by the two species, is an almost certain indication of their
very close relationship.
Parkinson reported
(1923, p. 108) that the trunk of this species measures 2 to 3 feet (61 to 91
cm) in girth at breast height (19.3 to 29.1 cm diam.), an unusual girth for an
Atalantia. This Andaman Island form (which Parkinson called
A. monophylla) has globose fruits 2.5 cm in diameter; it is common
everywhere in deciduous forests.
Engler (1931,
p. 328) recognized A. macrophylla as a good species distinct from A.
monophylla. Hochreutiner (1904, pp. 50-51) considered the form
from Bangka Island as a variety of A. monophylla differing "in having
somewhat larger leaves, less hairy twigs and especially in having a larger
calyx with convex acute lobes; in the typical A. monophylla the calyx
lobes are smaller and truncate-scarious." A specimen examined in
the Botanic Garden at Buitenzorg (now Bogor), Java, was a round-topped tree, 12
m high, with a deeply furrowed trunk 1.8 m in circumference (59 cm diam.),
forking at 1 m from the ground and branching profusely at 2 to 3 m from the
ground. No information is given by Hochreutiner concerning the size
of the fruits or the number of ovules in each locule of the
ovary.
Atalantia macrophylla is the
only species of the subgenus Atalantia that is found south of the Asiatic
mainland; it seems to have spread westward to the Andamans and southward to
Bangka from extreme southern Burma, retaining its large size and exceptional
girth, but showing diminished (although still large) fruit size.
The typical form of the species, growing in Tenasserim, has the largest fruits
of any of the Near-Citrus Fruit Trees (included in the genera Citropsis
and Atalantia).
3.
Atalantia racemosa Wight,15 Hook. Jour. Bot. 1:64,
pl. 122. Jan., 1834; A. racemosa Wt. & Arn. Prodr. 1:91. Apr.,
1834, nomen semi-nudum. Sclerostylis racemosa Wt. 1838?; S.
parvifolia Wt. 1838?; Lampetia racemosa (Wt.) Roem. Syn. Hesper.
1:42. 1846. Illus. Wight, loc. cit. pl. 122; idem,
Ill. Ind. Bot. 1:pl. 71. 1838?; idem, Icon. Pl. Ind. Or.
1:pl. 71. 1840; Beddome, Fl. Sylv. Anal. Gen. xlvii, pl. 7. 1871; Talbot,
For. Fl. Bomb. Sind. 1:p. 202, fig. 123. 1909; Swingle, Jour. Arnold
Arbor. 21:133, pl. 4, fig. 5. 1940.
Type.—Southwestern British India, "Madera…Alpine country"
(Wight). Herb. Univ. Glasgow.
Distribution.—Southwestern India: Konkan Coast to Travancore;
Ceylon. It is much more common in the Bombay Presidency than A.
monophylla, which is there rare (fide Cooke, 1903, p.
187).
Common name.—Bombay
atalantia.
This species was described by
Hooker (1875, vol. 1, p. 512) as follows: "Leaflet oblong-elliptic or
ovate-oblong, tip obtuse or 2-lobed, flowers racemed, rarely fascicled,
racemes pubescent or glabrous, calyx 4-lobed, lobes acute or obtuse, filaments
more or less connate, ovary 2-3 celled, ovules 2 collateral.…A small tree, armed
or unarmed, very similar to A. monophylla, but the flowers, instead of
being long-pediceled in short racemes or fascicles, are shortly-pediceled on
racemes sometimes 3 in. long, but at others almost reduced to corymbs; the
leaves are usually larger, and often broader, though quite small in Wight's
figure; the flowers are of the same size, but often very crowded and the buds
very globose, they are usually 4- but sometimes 3-5-merous, and the filaments
are free or variously united; the ovary is sessile on a small disk.—The great
distinction between these species is in the more or less regularly 4- (3-5-)
lobed calyx of this, the lobes of which are acute or obtuse. Fruit
globose, 3/4 in. [19 mm] diam. One of Wight's specimens has ovate
leaves, cordate at the base. Wallich’s 6358-E, from Penang (G.
Porter), in fruit only, with leaves cordate at the base, may be this or the
following [A. ceylanica], or something different from both."
This species, like A. monophylla, has the
stamens fused into a tube. In A. racemosa, however, the
stamens are often fused all the way to the tips into a cup-like cylinder
surrounding the ovary and the anthers are attached directly to the edge of the
cup, whereas in A. monophylla the tips of the stamens are free and only
the bases (about two-thirds to three-fourths of their length) are fused
together into a tube which is somewhat irregular because the free tips vary in
length. The flower buds of A. racemosa are globose and the
calyx differentiates into four or five (rarely three) lobes, whereas in A.
monophylla the flower buds are oblong or ovate, pointed at the tips, and the
calyx shows no division into sepals but splits more or less irregularly into
two or three lips often to the base on one side as the bud opens.
The fruits are much like those of A. monophylla. They are
globose, about 18 to 20 mm in diameter, and show the three to five lobes of the
persistent calyx. The flesh is composed of pulp-vesicles.
3a. Atalantia racemosa
var. henryi Swing. Jour. Arnold Arbor. 21:127. 1940.
Illus. Swingle, loc. cit. pl. 4, figs.
1-4.
Type.—China, Yunnan Province,
Szemao, altitude 1,220 m (Henry, No. 12930). in [sic] Herb.
Arnold Arbor.; photographs and serial microtome slides, S. and T. Nos. 660-A,
660-B, 660-C, and 660-D (14 slide [sic] with 464 cross sections and 156
longitudinal sections). in [sic] Herb. Natl.
Arbor. Paratype: Same collection, in Natl. Herb., sheet No.
459320.
Distribution.—China: Szemao and
Chi-li district in southern Yunnan Province near the Indo-Chinese border at
altitude 1,100-1,800 m.
Common
name.—Henry's atalantia.
A small tree, 4-7
m high, ultimate branches slender, 2-3 mm diam., soon terete, spineless; leaves
glabrous, lanceolate, short-acuminate at apex, acumen 5-8 X 4-3 mm, tip blunt or
even slightly emarginate, cuneate at base, lateral veins very numerous, 15-30
on each side, not all clearly marked, arising at angles of 50°-70° with the
midrib, margins subentire, petioles 5-10 X 1.2-2 mm, glabrous, coriaceous, more
or less wrinkled, with a deep narrow channel, 0.5-0.8 mm wide on upper side,
articulated with the leaf blade; inflorescences single axillary racemes, 1-2 cm
long, with 5-15 flowers borne on slender pedicels 3-8 mm long, peduncles,
pedicels, and calyx pubescent, flower buds subglobose, 3-4 mm diam., calyx
lobes rounded, 1.8-2 mm long, 2-3 mm wide, margins thin and ciliate, petals
oblong, rounded at apex, stamens 8, filaments glabrous or very sparingly short
ciliate, cohering irregularly in groups, sometimes almost to the tips, in other
flowers free almost to base, anthers about 2 mm long, attached near the middle
to the narrowed filament, connective bearing 1 medium-sized oil gland near
tip; disk cup-shaped, 0.5 mm high, 2 mm wide, pistil 3.5-4 mm long, ovary ovate,
1.5 X 1.5 mm, 4-locular with 2 ovules in each locule, with 1 medium-sized oil
gland at top of each locule, style short and thick, 2 mm long (including
stigma), 0.7-0.9 mm wide, stigma not clearly distinguished from the style,
about 0.9-1 mm wide, with 4 stylar canals with 2 medium-sized oil glands in
each space between two stylar canals; fruits subglobose, 1.5-2 mm diam., borne
sparingly (1-2?) on each raceme on pubescent pedicels 3-5 X 1.5-2 mm, with
numerous sessile pulp-vesicles (scanty in ripe fruit from seed pressure?),
seeds 1-3, ovoid-oblong, 12 X 7 mm, monoembryonic.
This variety resembles the typical A. racemosa
of southwestern peninsular India and Ceylon in its racemose flower
clusters, but it has larger leaves, flowers with longer pedicels and usually
with a 4-locular ovary (not 2- or 3-locular) and with the staminal filaments
rarely completely connate into a long tube.
It
is remarkable that so far as is known neither this variety nor the species
itself has been found in the vast region, some 2,700 kilometers across, that
separates the Mekong River valley of southern Yunnan from the Western Ghats in
Bombay State, India. There are many instances of discontinuous
distribution of species or of varieties of a single species belonging to the
tribe Citreae but few are as striking as this one.
4. Atalantia citroides Pierre
ex Guill. in Lecomte, Not. Syst. 1:178. 1910. Illus. Guillaumin,
loc. cit. p. 176, fig. 8(2); p. 177, fig. 9(3, 4); idem, in
Lecomte, Fl. Gén. Indo-Chine 1:pl. 24, figs. C and 5, 6. 1911; fig.
3-30 this work.
Type.—French
Indo-China, Cochin China, Mount Dinh (Pierre, No. 4011). Herb. Mus.
Hist. Nat., Paris.
Distribution.—South
Vietnam: Cochin China; Cambodia.
Common
name.—Cochin China atalantia.
Guillaumin's
original description of this species reads, in translation, as follows: "Trees
5-10 m high, glabrous, spines straight, few, 5 mm long, in the axils of the
leaves; branches grayish. Petioles glabrous, 1 cm long.
Leaves coriaceous, ovate (5-9 X 2-4 cm), entire, attenuate at the base,
attenuate and emarginate at the apex, primary and secondary veins prominent
above and below, oil glands visible only by being pellucid [to transmitted
light]. Flowers very glabrous, in the axils of the leaves, pedicels
glabrous, 1 cm long, with bracts at the base; calyx cupulate, with 3 or 4 lobes,
obtuse at the apex; petals 4, reflexed, ovate, attenuate at the apex; stamens
10, almost as long as the petals, more or less equal, connate for 2/3 of their
length, anthers ovate, apiculate; disk annular at the base of the ovary; ovary
ovoid-elongate, terminating in an articulated cylindric style, caducous at
maturity, stigma capitate, locules 3-5, ovules 1-2 [in each locule],
collateral. Fruit very similar to a small orange, 2 cm diam., pulp
made up of succulent vesicles, seeds ellipsoid, about 1 cm long."
The specimens from Cambodia were said by
Guillaumin to differ from those from Cochin China in having the young twigs,
as well as the peduncles and pedicels of the flower clusters, slightly
pubescent, as shown in his figure C, cited above.
This species has the filaments of the stamens
united into a cylindrical tube like A. monophylla and A.
racemosa. The tips of the filaments (about one-third of their
length) are free and the anthers are oval and apiculate, not triangular, as in
A. monophylla. The petals are narrowed at the tip, not
broadly rounded. The inflorescences resemble those of A.
monophylla, having rather long-pediceled flowers clustered on short
inflorescences in the axils of the leaves. The calyx is much like
that of A. racemosa, with three to four sepals. The fruits
resemble small oranges about 20 mm in diameter, with a rough peel and the flesh
formed of sessile, conical pulp-vesicles. The seeds are ellipsoid,
about 10 mm long.
This species grows to be a
medium-sized tree, 5 to 10 m high, and is probably larger and more vigorous
than any other species of the subgenus Atalantia except A.
macrophylla.
Although this species looks
much like A. monophylla and has flowers with similar staminal tubes, it
differs greatly in having a calyx with distinct sepals, unlike the curious
balloon-like calyx of A. monophylla and A. macrophylla that
splits irregularly into two or three lobes as the flower bud
expands. Tanaka (1930a, p. 163) made this species a synonym
of A. monophylla, overlooking this important difference.
Guillaumin, who studied A. monophylla, also widely distributed in
Indo-China, considered it to be clearly distinct from A. citroides.
5. Atalantia wightii Tan. Bul.
Soc. Bot. France 75:714. 1928.
Type.—Nilgiri Hills, Madras, southern British India (Bentham, No.
1531). Herb. Kew.
Distribution.—India; northern Canara, Bombay; Mysore and Madras (fide
Tanaka, 1937, p. 234).
Common
name.—Nilgiri atalantia.
Tanaka's original
description reads, translated, as follows: "Leaves oblong-ovate, coriaceous,
acute at the tips, rounded at the base, never attenuate; [stipuloid] paraphylls
linear, very short; racemes few-flowered; calyx lobes very narrow and acute,
glandular; stamens free (at length sometimes crowded); style evenly cylindric;
stigma obtuse." A typical specimen collected in Nilgiri Hills,
India (Bentham, No. 1531), preserved in the Rijks Herbarium at Leiden, Holland,
of which Tanaka provided Swingle a photograph, shows leaves 6-10.5 X 2-4.5 cm
(exclusive of petioles), acute at the tip and rounded or subcuneate at the
base, lateral veins numerous, 15-20 or more pairs arising at a wide angle
(60°-70°) with the midrib, margins entire; petioles very short, 4-6 X 1-2 mm,
wingless. Another specimen of Bentham's type collection (No. 1531),
preserved in the herbarium of the British Museum, London, shows the flowers in
axillary racemes or corymbs, with 8-10 or more very small flowers borne on
slender pedicels 4-5 mm long.
A specimen with
good flowers and young fruits in the herbarium of the Arnold Arboretum at
Jamaica Plain, collected by Father Anglade in southern India in the Kodaikanal
region, Pulney [or Pulni] Hills about 75 kilometers northwest of Madura,
identified by Tanaka ("Det. A-382") as Atalantia wightii, has smaller
leaves than the type specimens, blunt at the apex and emarginate, with the leaf
blades 4-7 (or 8?) X 1.5-3.8 cm, with fewer lateral veins (11-18); petioles 3-6
mm long; flowers small, densely crowded in axillary racemes, pedicels very
slender, 3-4 mm long; calyx lobes subacute at apex; petals linear, tapering to
an acute apex, 5.5-6.5 X 2.5-2.8 mm.
In
properly restored material from this specimen cut into serial sections the
floral organs appear as follows: Stamens 4.5-5 mm long, filaments 2.8-3.2 mm
long, 0.1 mm wide at base, tapering upward, 0.25-0.3 mm thick, cohering in
groups near the base, anthers without an oil gland in the connective; pistil
(including disk) 4.5-5 mm long; disk short, shallow-cupulate, 0.5-0.6 mm high,
1.5 mm wide; ovary ovoid, 2 mm long, 1.6 mm wide, merging into the thick style,
which is about 2 mm long, 1 mm wide, and merges into a stigma of nearly the same
width, slightly 2-lobed for about 0.4-0.5 mm at the top; young fruits fusiform,
8-9 X 5-5.5 mm (when dry), in restored microtome cross section 6-7 mm diam.,
2-locular with 2 collateral ovules in each locule, and a few pulp-vesicles
developing in the locule walls.
It seems
probable that A. wightii is somewhat related to A. simplicifolia
and A. roxburghiana. Like these two species, it needs
more study of the flower and fruit characters.
6. Atalantia simplicifolia (Roxb.) Engl. Die Nat.
Pflanzenfam. 3(4):192. 1896. Amyris simplicifolia Roxb. Fl. Ind.
2:244. 1832; Atalantia caudata Hook. f. Fl. Brit. Ind.
1:513. 1875.
Type.—"Pulo
Penang," Malaya? (coll.?). Herb. Brit. Mus.,
London.
Distribution.—Northeastern
India: Khasi Hills; also, fide Tanaka (1930b, p. 232), in
[northern] Burma but not yet found in Malaya or even in southern Burma
(Tenasserim).
Common name.—Khasi Hills
atalantia.
Hooker's original description of
A. caudata, collected by Hooker and Thompson at Churra, altitude 1,220 m
[4,000 ft.], Khasi Mountains, northeastern India, now considered a synonym of
A. simplicifolia, reads as follows: "Leaflet elliptic-lanceolate
caudate-acuminate with a broad 2-lobed tip; flowers small in very short glabrous
racemes, calyx very small 4-5 lobed, ovary 2-celled, ovules 2 collateral…A
glabrous unarmed shrub, with slender branches. Leaflets 3-4 in.
[7.5-10 cm], much narrower and more narrowed at each end than in any congener,
margin quite entire, tip sometimes dilated. Racemes 1/2 in. [13
mm], peduncles and pedicels much more slender than its congeners.
Flowers 1/4-1/3 in. [6-8 mm] diam., white, fragrant, usually 4-merous, buds
rather longer than broad. Calyx very small. Petals
obovate-oblong. Stamens quite free; anthers ovoid. Ovary
sessile on a disk narrower than itself; stigma subcapitate.—A very distinct
species, distributed as A. Roxburghiana? by T. Thompson and myself."
The type specimen of Roxburgh's Amyris
simplicifolia, preserved in the herbarium of the British Museum, shows the
following characters: leaves elliptical-lanceolate, 6.5-9.5 cm long (excluding
petioles), 2-3.6 cm wide, acuminate at the apex, with an acumen 6-8 X 4-1 mm,
the tip blunt and minutely emarginate, cuneate at the base and narrowed toward
the petiole; petioles 5-6 X 11.5 mm with a narrow (nearly closed?) canal on the
upper side, articulated with the blade, primary veins 10-16 with about as many
fainter ones between, arising at angles of 50°-60° with the midrib;
inflorescences axillary, peduncles 1-3 cm long, 0.4-0.6 mm wide, pedicles
almost all fallen (about 3.5 mm long?).
The
type specimen of A. caudata preserved in the Kew Herbarium looks very
much like-the type of Amyris simplicifolia except that the leaves are
slightly narrower, 2-2.8 cm wide, and have a longer acumen, 8-11 X 4-2 mm, with
a blunter, more deeply emarginate apex.
This
species is still incompletely known and in particular the flowers and fruits
are not adequately described. It is without doubt related to A.
roxburghiana but seems to be specifically distinct. It has been
found so far only in the Khasi Hills in Assam and in near-by parts of
northwestern Burma, more than 2,500 kilometers distant from the type locality
of A. roxburghiana at Malacca; it has not yet been found in southern
Burma (Tenasserim) about 1,800 kilometers distant.
The nomenclature of this species is
confused. The type specimen in the British Museum is credited to
Pulo Penang, but the species has never been found anywhere near
Penang. The original description of Amyris simplicifolia
Roxb. agrees fairly well with the type specimen, but a drawing of the species,
made by Roxburgh and published later by Wight (1840, pl. 72) under the name
Sclerostylis roxburghii, does not agree either with the description or
with the type specimen, as it shows short petioles not articulated with the
blade, the margins of which are decurrent well down the petiole. It
also shows only a few (five to seven) main lateral veins and about as many more
fainter and shorter ones, whereas the type specimen shows many more veins, both
strong and faint.
If the figure was made from
some other plant, not this species, and if the type specimen came, not from
Penang, but from Assam in northeastern India, or from northwestern Burma, then
undoubtedly the name A. simplicifolia will hold.
7. Atalantia roxburghiana
Hook. f. Fl. Brit. Ind. 1:515. 1875.
Type.—Malay Peninsula, Malacca (Griffith). Herb. Bot. Gard.,
Calcutta?
Distribution.—Malay
Peninsula, also Indo-China; fide Tanaka, varieties of this species also
occur in Thailand and southeastern China; also, fide Burkill (1931, p.
216), in northern Sumatra.
Common
name.—Malayan atalantia.
A sprawling
thornless shrub, ultimate branchlets slender, 1.5-2.5 mm diam., soon terete,
glabrous; leaves large, lanceolate or long-elliptical, tapering toward both
ends, but often acute or shortly subacuminate at apex and sometimes broadly
cuneate or rounded at base, 9-21 cm long (including petiole), 2-4 cm wide,
glabrous, subcoriaceous, with abundant veins visible on both surfaces, primary
veins very abundant, 10-12 stronger ones and as many fainter ones arising at an
angle of 70°-80° with the midrib; petioles short, 6-10 mm long, 1.3-1.8 mm
wide, with a very narrow, almost closed canal, 0.3-0.7 mm wide, on the upper
side, glabrous, articulated with the leaf blade; flowers not seen;
inflorescences (bearing fruits) axillary, 1-6 cm long, peduncle often simple,
bearing pedicels 5-6 mm long, 0.6-1 mm wide; calyx lobes 4, persistent,
glabrous, rounded, margins thin, sparingly ciliate; fruits subglobose, 1.5-2 cm
diam., young fruits 2-locular, with abundant sessile, conical pulp-vesicles,
older fruits often nearly full of oblong seeds, 10-12 X 4-6 mm; oil glands in
peel rather sparse and often large, 0.3-1 mm diam.
This seems to be a very well-defined species,
easily recognized by its being spineless and having unusually long leaves with
short petioles, less than one-tenth as long as the blade, which have a nearly
closed canal on the upper side. It is closely related to A.
simplicifolia, which has smaller acuminate or caudate leaves.
Two varieties of A. roxburghiana are described below.
The nomenclature of this species and of A.
simplicifolia is badly confused, so badly confused that it cannot be settled
finally without further study of the type material of both A.
roxburghiana and A. simplicifolia. It is possible that
the name here used can be considered as legal since it was based, in part, on
fruiting material that doubtless belongs to the species as here
described. Unfortunately, the original description was also based
on a misleading illustration of Amyris simplicifolia Roxb. which does not
agree with the original description of Amyris simplicifolia or with the
type specimen of it preserved in the British Museum.
7a. Atalantia
roxburghiana var. kwangtungensis (Merr.) Swing. Jour. Arnold Arbor.
21:129. 1940. Atalantia kwangtungensis Merr. Phil. Jour. Sci.
21:496. 1922.
Type.—China,
Kwangtung Province, Tung Sing (Ts'oong, No. 1936). Herb. Bur. Sci.,
Manila.
Distribution.—Known only from
the type locality.
Common
name.—Kwangtung atalantia.
The original
description, in English, reads as follows: "A glabrous shrub, the branches
terete, the branchlets somewhat compressed, unarmed. Leaves
subcoriaceous, olivaceous, oblong-elliptic, 10 to 14 cm long, 4 to 6 cm wide,
subequally narrowed to the acute base and to the acute or somewhat obtusely
acuminate apex, the upper surfaces slightly shining, the lower somewhat paler;
primary lateral nerves about 14 on each side of the midrib, prominent on the
lower surface, the ultimate reticulations rather close and distinct on both
surfaces, the glands invisible except by transmitted light; petioles about 5 mm
long. Inflorescences axillary, very short, of solitary or
fascicle-like racemes, the rachis 3 mm long or less. Flowers white,
about 6 mm long, their pedicels stout, 1 mm in length; bracteoles ovate,
obtuse, about 1 mm long, the margins ciliate. Calyx lobes 5,
orbicular, rounded, 1.5 to 2 mm wide, the margins slightly ciliate.
Petals 5, 4 to 5 mm long, oblong-elliptic, rounded, glabrous.
Stamens usually 8, the filaments about 3 mm long, flattened, glabrous, and
united for the lower one-half; anthers 1.2 mm long. Ovary glabrous;
style 1 mm long."
This variety differs from
the species in having usually 5-merous flowers (instead of 4-merous) and
staminal filaments cohering from the base for about half their length instead
of being free. Tanaka (1930b, p. 232) referred A.
kwangtungensis to A. roxburghiana and Merrill (1931, p. 311) accepted
this reduction. In view of the presence of characters in A.
kwangtungensis not yet found in typical A. roxburghiana, it appears
possible, even probable, that the southern Chinese form deserves to be
recognized as a variety, as done here, pending further study of the A.
roxburghiana complex of forms.
7b. Atalantia roxburghiana var. kerrii Swing.
Jour. Arnold Arbor. 21:129. 1940.
Type.—Thailand, Sam Roi Jawt (Kerr, No. 10943). Herb. Univ.
Aberdeen; photographs, fragments, and 12 serial microtome cross sections of a
fruit, S. and T. No. 684-A, slides 1-4, in Herb. Natl. Arbor., Washington,
D.C.
Distribution.—Known only from the
type locality.
Common name.—Kerr's
atalantia.
A small tree up to 5 m tall,
ultimate branches at first green and slightly angular, soon brownish, terete
and faintly striate longitudinally; internodes 1.2 cm long; leaves thick and
leathery, broadly oval or elliptical, smaller ones lanceolate, 9-15 cm long
(including the petiole), 3-8 cm wide (usually 6-8 cm), tapering to a blunt apex,
broadly cuneate at base with numerous small oil glands scattered over the whole
surface of the leaf blade but scanty near the margins which are entire or
faintly crenulate and slightly thickened at the very edge, lateral veins 7-12
pairs, nearly straight or slightly curved, forked at 5/6-6/7 of the distance to
the margin, making angles of 45°-60° (rarely 65°-70°) with the midrib, with
numerous small lateral veinlets that anastomose; petioles stiff, glabrous,
wrinkled, 8-13 mm long, with a shallow channel on the upper side, 1-2 mm wide,
0.4-0.5 mm deep; inflorescences axillary, paniculate (?), 2-10 cm long,
peduncle 1-2 mm diam., pedicels 5-8 X 1.25-1.8 mm; fruits subglobose, 1.5-1.8
cm diam., peel covered with large, slightly sunken oil glands, 0.5-0.9 mm
diam., 3-locular, pulp-vesicles numerous, sessile, 3-5 mm long, filling all
the space not occupied by the seeds; seeds ovoid 11.5-12 X 10-11 X 6-8 mm, with
a very thin, papery testa (which swells and separates from the embryo in hot
water), monoembryonic.
This remarkable
Atalantia has large, broad, coriaceous leaves; unfortunately it is known
only in the fruiting stage. It differs strikingly in its leaf
characters, not only from A. roxburghiana, but also from the other
species of Atalantia known from the Indo-Chinese region. As
the species of this genus are, most of them, variable in many of their
characters, it seems best to consider this striking form as a variety of A.
roxburghiana Hook. f., not uncommon in the Malay Peninsula and reported from
Indo-China, at least until flowers can be secured for study. This
variety is named in honor of Dr. A. F. G. Kerr, who did so much to make known
the rich flora of Thailand.
Subgenus Rissoa
Subgenus
Rissoa (Arn.) Swing. in Bailey, Stand. Cycl. Hort. 1:426. 1914.
Rissoa Arn. Nov. Act. Acad. Caes. Leop.-Carol. 18:324.
1836.
Type.—Rissoa ceylanica
Arn. = Atalantia ceylanica (Arn.) Oliv.
Stamens free; fruits with the locules nearly filled with large seeds, with only
a few sessile pulp-vesicles; leaves often accompanied at the base with a pair
of small stipuloid paraphylls.
The species of
this subgenus are very like those of the subgenus Atalantia in general
appearance but differ from them in having free stamens, and nearly dry fruits
with large seeds, almost filling the locules. Only two species of
Atalantia (A. ceylanica and A. rotundifolia) are now known
to belong to this subgenus, but probably several others will be placed here
when their fruit structure is better known.
8. Atalantia ceylanica (Arn.) Oliv. Jour. Linn. Soc. Bot.
5(Suppl. 2):25. 1861. Rissoa ceylanica Arn. Nov. Act. Acad. Caes.
Leop.-Carol. 18:324. 1836; Limonia monophylla L. Mant. Pl. Alt.
237. 1771; Sclerostylis ceylanica Wt. 1840; S. arnottiana Wt.
1840. Illus. Swingle, in Bailey, Stand. Cycl. Hort. 1:426, fig.
434. 1914 ; fig. 3-31 this work.
Type.—Ceylon (Wight No. ?), Glasgow Univ. Herb. (fide Tanaka,
1930b, p. 232).
Distribution.—Ceylon, low country; southern
India.
Common name.—Ceylon
atalantia.
This species was described as it
occurs in Ceylon by Trimen (1893, p. 227) as follows: "A much-branched bush,
usually armed with very sharp, stout spines, 1/4-1 in. [6-25 mm] long, bark
smooth, brown, young parts glabrous, l[eaves] very variable, 1 1/4-3 in. [3-7.6
cm long], lanceolate or oval-oblong or somewhat ovate, rounded at base,
emarginate or 2-lobed at apex, entire, coriaceous, veiny, petiole short, thick,
glabrous; fl[owers] 1/2 in. [13 mm diam.] or rather more, on somewhat slender,
glabrous ped[icels], usually crowded in short, very finely pubescent,
corymbose, or racemose cymes much shorter than the l[eaves], bracts minute;
cal[yx] lobes broad, shallow, often obscure, finely ciliate; pet[als] 4 (rarely
3 or 5), oval, with a broad claw, obtuse, thick, slightly concave, glandular;
stam[ens] 8, alternate ones longer, quite distinct, fil[aments] flat, rather
wider than anth[ers], anth[ers] cordate-ovate, apiculate; ov[ary] sessile,
smooth, 2-celled, with 2 collateral ovules in each cell; style short, stout,
stigma clavate; berry 1/2-3/4 in. [13-19 mm diam.], globular-ovoid,
2-4-seeded."
Hooker (1875, p. 511) described
it as follows: "leaflet obtuse, notched or 2-lobed, rarely obtusely acuminate,
flowers very shortly racemed, calyx 4-lobed, filaments free or 2 or 3 of them
more or less combined, ovary 2-celled, ovules 2-collateral." He
mentioned in the notes that the ovary is "sunk in the annular
disk."
Atalantia ceylanica is
remarkable for its nearly dry fruits, with the very few pulp-vesicles almost
crowded out by the large, plump seeds that almost completely fill the
locules. Small stipuloid paraphylls, 4 to 6 mm long, often occur
singly or in pairs at the base of the petiole. This species seems
to be related to A. guillaumini, from Indo-China, which apparently has
lost its pulp-vesicles and has still larger seeds that completely fill the
locules of the fruit.
9.
Atalantia rotundifolia (Thwaites) Tan. Bul. Soc. Bot. France
75:714. 1928. Sclerostylis rotundifolia Thwaites, Enum. Pl. Zeyl.
46. 1858; Atalantia ceylanica var. rotundifolia (Thwaites) Oliv.
1861.
Type.—Ceylon, Muratte (Thwaites,
No. 3295). Herb. Kew; photographs, fragments, and serial microtome
sections of type material, S. and T. Nos. 652-A, 652-B, 652-C, and 682-D, 17
slides, in Herb. Natl. Arbor., Washington,
D.C.
Distribution.—Ceylon: "Montane
zone at about 4,000 ft. [1,220 m] and upwards"; also in Madras, India, and
eastward in Indo-China (Vietnam), Annam and Cochin China (fide Tanaka,
1928b, p. 714).
Common
name.—Dwarf Ceylon atalantia.
Thwaites's
original description reads (translated with the addition of the English
account) as follows: "Spineless; leaves rounded or ovate-rounded, retuse,
coriaceous; inflorescences racemose, grouped at the ends of the
branches. A small tree, very much branched, glabrous.
Leaves 0.75-1.75 in. [19-45 mm] long, 0.5-1.4 in. [12-39 mm] wide; petiole 1-2
lin. [2-4 mm] long. Racemes axillary and terminal, solitary, or in
clusters of 2 or 3, 5-10-flowered, as long as the leaves. Flowers
4-merous. Calyx profoundly 4-lobed; lobes rounded.
Ovary oblong, short-stalked, attenuate into a cylindric style of equal length
ending in a compressed-dilate top with stigmatic margins; locules 2, 2-ovulate
[Latin to here].
"Closely allied to the
preceding species [= A. ceylanica], but distinguished at once by the
absence (apparently) of spines, and the different shape of the
leaves. The calyx is deeply four-lobed, whereas in S.
[Atalantia] ceylanica it is scarcely more than undulated; the
style is also more slender in the present species, and the ovary more decidedly
stalked. In both species the latter organ is surrounded at its base
by a free, glandular annulus."
A photograph
of Thwaite's [sic] type, No. C.P. 3295, in the Rijks Herbarium at
Leiden, sent Swingle by Tanaka, shows that this species has the leaves densely
crowded on the twigs, because of the shortness of the internodes.
No spines can be seen. Many of the leaves are elliptical in shape,
abruptly rounded at the narrow base but usually emarginate (often rather
deeply) at the apex, giving the appearance of an obcordate leaf that had been
greatly compressed laterally. The leaves appear to be tufted at the
ends of the short twigs, where the internodes must be very short, probably only
4 to 6 mm long in some instances.
The whole
appearance of this plant is reminiscent of the spineless or nearly spineless,
short-internoded mutations of Severinia buxifolia—for example, the brachytic form—and the myrtle-leaved mutation of the sour or
Seville orange, Citrus aurantium var. myrtifolia. Possibly this species arose from
A. ceylanica as such a brachytic, spineless mutation which in time bred
true and persisted at higher altitudes. Atalantia rotundifolia
therefore is probably to be considered as a satellite species of A.
ceylanica. It is possible that A. rotundifolia will some
day be used for hedges or other ornamental purposes in moist subtropical
regions.
SPECIES OF UNCERTAIN RELATIONSHIPS
10. Atalantia guillaumini Swing. in Lecomte, Not. Syst.
2:159. 1911. Atalantia disticha Guill. (non Merr.) in Lecomte, Fl.
Gén. Indo-Chine 1:673. 1911. Illus. Swingle, loc. cit.
2:162. fig. 1.
Type.—French
Indo-China, southern Tonkin, Lang-hé, on Mount Dên (Bon, No. 4047).
Herb. Mus. Hist. Nat., Paris.
Distribution.—North Vietnam: known only from southern Tonkin, from Mount
Dên, Mount Nam-cong, Mount Luong-xa, and Mount Thin-chan (all from collections
made by H. Bon.)
Common name.—Tonkin
atalantia.
Swingle's original description,
translated, reads as follows: "Leaves ovate-lanceolate, apex obtuse, base
rounded or deltoid with many lateral veins running parallel [margins entire];
petioles articulated at both ends, not winged, not [deeply] channeled; calyx
persistent, sepals 4-5, subtriangular, with sparsely ciliate margins; fruits
solitary, 2.4-2.8 cm diam., globose, with 3 or 4 segments, segments 2- (or 1-
?) seeded, destitute of pulp-vesicles; seeds large, with membranous
testa." [Latin diagnosis to here; continuing, in French, the
translation reads:] "A small tree, 3-4 m high, with spiny, pubescent
branches. Twigs more or less angular when very young, soon becoming
rounded and glabrous. [Fruiting twigs] spineless.
Internodes 1-2 cm long. Leaves large, oval or oval-lanceolate,
rounded at the tip and base; the small leaves lanceolate, more or less pointed
at the tip and base; leaf blades 55-85 mm long, 25-60 mm wide, always clearly
distinct from the petiole (which is, moreover, articulated at the point of
junction with the leaf blade), glabrous, without oil glands except at the
margins; lateral veins numerous, 20-30 [sometimes fewer], parallel and almost
straight for 3/4-5/6 of their length, confluent by anastomosis near the margin;
petioles 5-9 mm long, flattened above with the edges slightly elevated, brown
in dry specimens, with many or few very slender hairs; stipules [stipuloid
paraphylls] short, narrow, 2 mm long, 0.5 mm wide, veinless.
Inflorescences few-flowered, spicate, 3-4 cm long; axillary or else bearing
only isolated flowers or a single fruit in the axils of leaves near the tips of
the young twigs; flowers not seen; fruit pedicels 6-8 mm long, 1.5-2 mm
diam. Calyx persistent, composed of 4-5 distinct sepals that are
more or less triangular, sparsely ciliate on the margins. Fruits
solitary, spherical or slightly pointed, 2.4-2.8 cm diam. with 1-3-fertile
segments; seeds 2 in each segment, very large, 12-15 mm long, 8-9 mm wide,
rounded at both ends, more or less angular from mutual pressure, with a very
thin membranous testa; pulp-vesicles absent, skin glandular and rough, but
thin."
Guillaumin (1911, p. 673) described
this plant (under the name Atalantia disticha [Blanco] Merrill) as a
"tree 4 m high, spineless," but went on to say, "branches…with small erect
spines," and later on, "petioles…furnished with 2 erect stipules [stipuloid
paraphylls], in the form of spines reaching 5 mm [in length] or in the form of
minute linear leaf blades [paraphylls] showing a central vein and a few
veinlets."
This species much resembles A.
ceylanica (1) in having large fruits filled with very large seeds; (2) in
the general character of the leaves, which have few or no oil glands except
along the margins; and (3) in having frequently one or more paraphylls more or
less like stipules at the base of the petiole, usually more spinelike than in
A. ceylanica. The ripe fruits of A. guillaumini do not
show pulp-vesicles, or at least Swingle did not find any in the type
specimen. The ripe fruits of A. ceylanica usually show traces
of a few pulp-vesicles, although almost all of them are crowded out by the very
large seeds that almost completely fill the locules. The still
larger seeds of A. guillaumini may perhaps obliterate all the
pulp-vesicles in many of the fruits, or even in all of them. The
study of young fruits of this species should show whether a few pulp-vesicles
are present or whether they have been completely suppressed. If the
latter should prove to be true, a very interesting and important taxonomic
problem would be raised, since the possession of pulp-vesicles of a special
type is an important character of the genus
Atalantia.
Severinia disticha,
to which this species was assigned by Guillaumin (1911, p. 673), is a very
different plant, having small fruits with rudimentary, round, stalkless
pulp-vesicles crowded into the periphery of each locule. A single
seed of Atalantia guillaumini is larger than an entire fruit of
Severinia disticha. Severinia disticha has, moreover,
the cup-shaped disk enclosing the lower part of the ovary, as in other typical
species of Severinia.
11.
Atalantia hainanensis Merr. & Chun, ex Swingle, Jour. Arnold Arbor.
21:20. 1940. Illus. Swingle, loc. cit. pl. 4, figs.
3-7.
Type.—China, Hainan Island,
Po-Ting (How, No. 72807). Herb. Arnold Arbor, [sic] Harv.
Univ., Cambridge; photographs, fragments, and serial microtome sections, S. and
T. Nos. 261-A, 261-B, 261-C, 12 slides, in Herb. Natl. Arbor., Washington,
D.C. Cotype: Same locality (How, No. 73976), Herb. Arnold
Arbor.; serial microtome sections S. and T. Nos. 263-A and 263-B, in Herb.
Natl. Arbor.
Distribution.—Known also
from Yaichow, Hainan Island. Not uncommon, usually in forests near
streams at altitudes of 100-640 m.
Common
name.—Hainan atalantia.
A small spineless
shrub, 1-2 m high; young twigs slightly angled but soon becoming cylindrical;
leaves simple, persistent, thick and coriaceous, elliptical or broadly
elliptical, slightly acuminate or bluntly pointed at the tip, often
emarginate, cuneate at the base, narrowing gradually into the petiole, very
variable in size, usually 6-15 X 2.5-6 cm, but sometimes as large as 20 X 8 cm
including the petiole, margins entire or faintly undulate; petiole not
articulated with the leaf blade, 5-10 mm long, more or less pulvinoid, with a
channel on the upper side formed by the decurrent leaf margins; inflorescences
axillary, few-branched panicles, short (1-1.5 cm); pedicels slender, 2-3 X 1
mm; flower buds small, 3-3.5 mm long, 2-2.5 mm wide; calyx with 5 triangular
lobes, thickened, roughened with oil glands except at the margins, which are
thin and ciliate; petals 5, white; stamens 10, filaments flattened, connate at
the base, free above, anther with 1 large oil gland in the connective; ovary
small, 1.5 X 1 mm, ovoid, with 2 locules, each with 2 collateral ovules, top of
ovary with 1 large oil gland above each locule; style slender, 1.25 X 0.3-0.4
mm, stigma nearly isodiametric with the style, with 2 pairs of large oil
glands; disk cupulate, about 0.35 mm deep, glabrous, completely surrounding the
basal 2/5 of the ovary; fruits (young) ellipsoid, 8-10 X 4-5 mm, peel green,
with numerous oil glands, surmounted by the small persistent style, 1 X 0.6-0.7
mm; seeds 1 (or more?) to a fruit, large, ellipsoid, about 10 X 7 X 5 mm,
monoembryonic (?), cotyledons with numerous oil glands.
This curious plant, clearly a very distinct new
species, is hard to place in the absence of mature fruits. It has
leaves varying greatly in size (2.5 to 19.5 by 0.8 to 7.5 cm) which have much
the same general aspect as those of Severinia buxifolia, a species it
further resembles in having a cupulate disk and a 2-locular ovary with a large
oil gland over each locule. One specimen not considered in the leaf
measurements given above (How, No. 73818) from Po-Ting, Hainan, in the
herbarium of the Arnold Arboretum, shows relatively very small leaves, only 2.5
to 5.5 by 0.7 to 1.5 cm, narrowly elliptical, very blunt-pointed and retuse at
the tip. Young fruits with persistent calyces on this specimen
were, however, almost identical with those of the large-leaved specimens from
the same locality in Hainan. However, serial microtome sections made
of the immature fruits of the cotype (How, No. 73976) show the locule walls
lined not with a clear-cut inner layer of stalkless subglobose pulp-vesicles
but rather with large and variable-sized oil glands (?), not all in a definite
layer but with a few of them scattered between the much smaller oil glands of
the peel and the inner locule wall. Study of the fruits of this
species at all stages of development may show that these structures have a
merely superficial resemblance to and no true homology with the primitive
pulp-vesicles of Severinia buxifolia and other typical species of
Severinia. The typical species of Atalantia have
sessile, broad-based, conical pulp-vesicles growing out from the dorsal locule
walls which with the seeds fill the locules completely. However,
A. ceylanica, the type species of the subgenus Rissoa, shows in
the mature fruit very few pulp-vesicles, perhaps because the very large seeds
almost completely fill the locules. Atalantia hainanensis,
like the anomalous A. guillaumini, of which only the very large,
subglobose, fully mature fruits are known, does not seem to have any
pulp-vesicles. These two anomalous species of Atalantia need
critical study at all stages of the developing fruits in order to be certain of
their taxonomic placement.
GROUP C. THE TRUE CITRUS
FRUIT TREES
This group includes six
genera, Fortunella, Eremocitrus, Poncirus, Clymenia,
Microcitrus, and Citrus, all marked by having orange or lemon-like
fruits with highly specialized, slender-stalked, usually more or less fusiform
pulp-vesicles filling all the space in the segments of the fruit not occupied
by the seeds, and also by having at least four times as many stamens as
petals. All the genera have persistent unifoliolate or simple
leaves except the monotypic genus Poncirus, which has trifoliolate,
deciduous leaves. Clymenia differs in having simple leaves
borne on short, wingless petioles, with numerous, nearly straight veins
prominent on the lower surface, somewhat resembling the venation of the leaves
of Wenzelia and Monanthocitrus of the subtribe Triphasiinae.
All the genera in this group except
Clymenia have fusiform pulp-vesicles borne on very slender stalks,
sometimes short, sometimes long. In Clymenia the
pulp-vesicles are subglobose or pyriform in shape, usually having subglobose
apices and being narrowed into somewhat contracted bases; the bases, however,
are never reduced to slender stalks of variable length, as they are in the
pulp-vesicles of the other genera of this group. All the genera
except Clymenia have all, or at least the majority, of the pulp-vesicles
attached to the dorsal walls of the locules. A few species of
Citrus (subgenus Papeda) have a few or sometimes many
pulp-vesicles attached to the radial walls, but only on the distal portion of
these lateral walls nearest the periphery of the fruit.
Clymenia differs from all the species of the subgenus Citrus of
the genus Citrus in that many of the pulp-vesicles, probably a large
majority of them, are attached to the radial walls of the segments and occur on
these walls nearly to the center of the fruit (three-fourths to four-fifths of
the way). Although Clymenia has much simpler pulp-vesicles,
without specialized apices, the other fruit characters are very like those of
Citrus, to which it is obviously related.
The structure of the pulp-vesicles is of great
taxonomic importance in the classification of the genera of the subtribe
Citrinae. Found only in this subtribe, these unique structures reach
their highest development in group C, the True Citrus Fruit Trees.
In contrast with group B, the Near-Citrus Fruit Trees, in which the
pulp-vesicles are conical in shape, tapering into more or less acute apices and
having broad, sessile bases, the pulp-vesicles of the True Citrus Fruit Trees
are never conical in outline, but are fusiform, being narrowed at the base
(except in Clymenia) into slender stalks (varying considerably in length
in different pulp-vesicles of the same locule) and into a point at the
apex. In Clymenia the pulp-vesicles are different from any
others occurring in the True Citrus Fruit Trees in being pyriform and in not
being slender-stalked at the base nor having acute apices.
The pulp-vesicles of all the genera included in
this group, with the possible exception of Clymenia (not yet available
for study in the living condition), contain droplets of oil. These
oil droplets are very abundant in the pulp-vesicles of Poncirus,
Microcitrus, and the species of Citrus belonging to the subgenus
Papeda, but are fewer in the pulp-vesicles of Fortunella,
Eremocitrus, and the species of Citrus belonging to the subgenus
Citrus. Similar oil droplets (or even wax particles) occur
also in the pulp-vesicles of the Near-Citrus Fruit Trees, Citropsis and
Atalantia.
Three genera in group C,
Clymenia, Eremocitrus, and Poncirus, are
monotypic. Two other genera, Fortunella and
Microcitrus, have four and six species respectively. All five
of these genera have a limited area of distribution much more restricted than
that of Citrus, which is the dominant genus of the subtribe with the
largest number of species (sixteen are recognized in this chapter).
The region occupied by the six genera comprising
the True Citrus Fruit Trees is a long, barrel-shaped area about 9,000
kilometers (5,590 miles) long and 3,200 kilometers (1,990 miles) wide in the
center, tapering to about 2,600 kilometers (1,615 miles) at the
ends. This barrel-shaped area has its long axis slanting from the
northwest (northeastern India to north-central China) to the southeast
(east-central Australia to New Caledonia). At its broadest part the
area extends from Java to the eastern shores of the Philippine
Islands. If, as some experts believe, Citrus is native to
southern Japan, there would be an extension of the northern corner of this area
to include Citrus tachibana. Citrus is native to the whole of
the area except in the southernmost corner (northeastern Australia), where it is
replaced by Microcitrus and Eremocitrus, and in the extreme
northernmost corner (northern China), where it is replaced by
Poncirus. Fortunella occurs in southeastern China, where
Citrus is probably also native, and Clymenia occurs in the
Bismarck Archipelago (the extreme easternmost part of the area), where one
species of Citrus is probably also native.
The six genera of group C are all obviously
closely related and all (except the newer, little studied genus
Clymenia) have been grafted on one another and hybridized with one
another.
Fortunella resembles
Atalantia (a Near-Citrus Fruit Tree of group B) in having 3 to 5- (rarely
6- or 7-) locular ovaries with only two ovules in each locule, also small
fruits and thick, stiff leaves, but differs in having highly organized,
slender-stalked pulp-vesicles. Eremocitrus has ovary and
fruit characters much like Fortunella, but it has undergone striking
xerophytic adaptations that have culminated in the acquisition of leaves very
diverse in structure from those of the other genera of the True Citrus Fruit
Trees. Poncirus stands alone in group C in having
trifoliolate, deciduous leaves and winter buds well protected by bud
scales. It has pleiomerous ovaries with six to eight locules, with
many ovules in each locule. In these ovarial characters it agrees
with Microcitrus and Citrus. Poncirus undoubtedly
represents an ancient offshoot from the True Citrus Fruit Trees that pushed
north into China and in so doing acquired deciduous leaves and bud scales and
developed great resistance to winter cold. Clymenia is
probably the most primitive of all the genera comprising the True Citrus Fruit
Trees. Although it has fruits (said to be sweet and edible) that
appear much like those of Citrus, it differs widely from Citrus
in having more primitive pulp-vesicles and in having leaves that resemble those
of the genera Monanthocitrus and Wenzelia of the subtribe
Triphasiinae, the Minor Citroid Fruit Trees. Microcitrus is a
primitive genus related to Citrus; perhaps some of its species are very
like the ancestral species from which Citrus developed.
Of the six genera comprising the True Citrus
Fruit Trees, Fortunella, especially the subgenus Protocitrus, is
the most simple and primitive of the main branch that ends in
Citrus. Microcitrus represents a higher stage of
evolution and is still closer to Citrus. Eremocitrus
and Poncirus are genera profoundly modified, the one to withstand
semiarid climates and the other winter cold. Clymenia is the
Cinderella of the group; it shows some affinity with the Papeda subgenus
of Citrus and even some relationship to certain of the Primitive Citrus
Fruit Trees, namely, Pleiospermium, Limnocitrus, and
Burkillanthus, on the basis of which a new theory on the origin of the
True Citrus Fruit Trees is possible (see under Clymenia; also under Pleiospermium).
Four of
the six genera in group C are clearly adapted to fit exceptional climatic or
soil conditions, conditions under which Citrus grows poorly or fails
completely. Eremocitrus is a pronounced xerophyte, able to
grow in semiarid regions in northeastern Australia that are too dry to support
Citrus. Microcitrus is semi-xerophytic and can endure
long drouths. Poncirus has become deciduous and is able to
endure very severe cold in winter, a condition fatal to Citrus and even
to Fortunella. It also has developed well-protected flower
buds that form during the early summer and push into bloom from old twigs the
following spring. Fortunella can withstand long spells of
warm weather in winter or early spring without showing new growth, thereby
escaping all danger of injury from spring frosts, often very injurious to
Citrus. These four genera can grow where no species of
Citrus can thrive; perhaps all of them are in fact survivals that escaped
destructive competition with Citrus by occupying territory where the
latter genus could not live. The other relative of Citrus in
this group, Clymenia, is so little known as yet that it is not possible
to state whether it has any physiological or structural adaptations that permit
it to survive where Citrus would fail. Perhaps, like the
four other genera of limited geographic range, it will on further study show
adaptive structures and physiological peculiarities that will give it an
advantage over Citrus in the regions where it occurs in a wild state.
There is every indication that
Eremocitrus, Microcitrus, Poncirus, and Fortunella,
because of their ancient and deeply inbred adaptations to special climatic and
soil conditions, will prove important in breeding new types of citrus fruits and
new rootstocks able to resist disease and able to endure unfavorable climatic
and soil conditions that no Citrus species can withstand. It
is for this reason that every effort has been made to present as full an
account as possible of these close relatives of Citrus, all of which can
very probably be hybridized with Citrus and also with one
another.
XXI. Fortunella
Swingle
XXI. Fortunella
Swingle, Jour. Wash. Acad. Sci. 5:167.
1915.
Type species.—Citrus
margarita Lour. = Fortunella margarita (Lour.)
Swing.
Distribution.—Southern China,
probably now only in cultivation.
Common
name.—Kumquat.
Shrubs or small trees;
young branches angular, the older ones rounded; spines borne singly at one side
of the bud in the axils of the leaves, or wanting; leaves 1-foliolate, rather
thick, blunt-pointed or even retuse, acute or rounded at the base, veins
evident above, scarcely showing beneath, lower surface pale green, densely
glandular-dotted; petioles narrowly winged or merely margined, sometimes not
articulated with the leaf blade; flowers borne singly or in few-flowered
clusters in the axils of the leaves, hermaphrodite, 5-merous (rarely 3-, 4-, or
6-merous); flower buds, 8-10 mm long, more or less angular in cross-section;
petals 5 (rarely 4 or 6), white, acute, 8-12 mm long; stamens 16 or 20,
polyadelphous, cohering irregularly in bundles; filaments broad, but tapering
at the tip; pistil seated on a well-marked cylindrical disk; ovary subglobose,
with 3-7 locules, with 2 collateral ovules in each locule; ovary merging
gradually or abruptly into the short style, this usually shorter than the
ovary, sometimes shorter than the stigma; stigma capitate, symmetrical,
cavernous within because of the large, deep-seated oil glands (1/4-1/5 the
diam. of the stigma); fruits small, usually 2-4 times as long as the petiole,
ovoid or globose; peel rather thick, fleshy, aromatic, and sweet flavored,
containing large immersed oil glands; segments 3-7; pulp-vesicles small,
fusiform or subglobose, stalked, containing an acid juice; seeds ovate in
outline, smooth; embryo pistache green, germination with hypogeous cotyledons;
first foliage leaves broadly ovate, subsessile, opposite as in
Citrus.
The genus Fortunella
resembles Citrus in the general appearance of the stems, twigs, spines,
leaves, flowers, and fruits and in having the polyadelphous stamens cohering in
bundles and normally four times as numerous as the petals. It
differs from Citrus: (1) in having an isomerous or hypomerous ovary
normally with three to five, rarely six or seven, locules (not polymerous with
8 to 15 or more locules); (2) in having two collateral ovules in each locule
(not 4 to 12); (3) in having a cavernous stigma containing a few large, deeply
immersed, lysigenous oil glands, usually in pairs, oval in cross section, with
the radial diameter longer, about one-fourth to one-fifth the diameter of the
stigma (in Citrus the homologous oil glands are so much smaller
[one-tenth to one-fifteenth the diameter of the stigma in C. sinensis]
that they do not give a cavernous character to the stigma); (4) in having the
under surface of the leaves pale green, nearly veinless, and with very
numerous, small, deep-green glandular dots; (5) in having very small fruits with
acid pulp and a sweet, edible, more or less pulpy skin; (6) in having small,
more or less angular flower buds.
The kumquat
bears abundant orange- or flame-to-orange-colored fruits of small size, often
less than an inch in diameter. Fortunella margarita and F.
japonica, widely cultivated in China and Japan and in all subtropical
regions, have fruits with a relatively thick, fleshy, sweet, edible peel, with
four to seven segments filled with mildly acid pulp. A third
species, with long, slender leaves and long petioles, F. polyandra,
commonly cultivated in the Malay Peninsula, has larger globose fruits with a
thin peel. A fourth species, F. hindsii, which grows wild in
the mountains of southern China and on the island of Hong Kong, has very small
globose fruits with three to four segments.
The kumquat orange, though described by early Chinese writers on agriculture,
remained virtually unknown to Europeans until recent times. The
kumquat is mentioned in many early Chinese works and described in some detail
by Han Yen-chih (1923) in his treatise on the oranges written in
1178. Later works of both Chinese and Japanese authors treat of it
fully, often with fairly good illustrations.
The first vague description of the kumquat orange in European literature was
published by Ferrari in 1646 in his Hesperides and was based on reports
made to him by Alvaro Semedo, a Portuguese Jesuit who lived for twenty-two
years in China. Ferrari's successors, Sterbeeck, Volckamer, Risso
and Poiteau, and other authors of monumental illustrated works on citrus
fruits, have added nothing to our knowledge of the kumquat.
Full descriptions of the round and oval kumquats
were published by Hume (1903, 1909), but not until 1912 was there a good
account of these plants published in Europe, when Trabut (1912) described them
and distinguished them from the so-called "chinosis" or "chinotto" (Citrus
aurantium var. myrtifolia), with which they had been confused by
Volckamer (1708) and many subsequent European writers. Two years
later, Trabut (1914) published in Algeria a fuller illustrated account of these
plants and gave an explanation of the failure of the kumquat to become known
and be propagated in continental Europe, namely, that seedlings do not thrive
and, furthermore, that viable seed is hard to secure; moreover, he stated that
the kumquat, when grafted on sour or Seville oranges (the stock generally
used), did not succeed at all.
One of the
kumquats, F. polyandra, is native to tropical regions; the other three
grow in cool subtropical or even warm temperate regions. The type
species of Fortunella, F. margarita, and the closely related F.
japonica not only are resistant to cold when in a dormant condition but also
exhibit the highest degree of winter dormancy of any of the True Citrus Fruit
Trees. These two species of kumquats can endure fairly warm
weather in winter or early spring lasting for many days or even for some weeks
without starting new growth (Swingle, 1910, 1913e; Swingle and Robinson
1923, p. 229.)
Fortunella approaches
Atalantia, and differs strikingly from Citrus, in having only two
collateral ovules near the top of each locule (Citrus has 4 to 12
ovules). However, Fortunella differs from Atalantia in
having four times as many stamens as petals (instead of twice as many) and
agrees with Citrus in having similar twig, leaf, spine, flower, and fruit
characters. The leaves, however, show very many more oil glands on
the underside than are present in any species of Citrus, often ten times
as many. In many superficial fruit characters Fortunella
agrees with the Australian desert lime, Eremocitrus glauca.
The seeds are very different, however, and the stem, twig, leaf, and flower
characters are so strikingly different that it is not possible to regard these
genera as being very closely related.
THE SPECIES AND HYBRID PROBLEM IN FORTUNELLA
The species of Fortunella, in spite of
their small number, have been very inadequately studied, largely because, until
recently, only one species was cultivated in either Europe or the United States
and the others were represented only by scanty herbarium material in a few of
the largest herbaria.
The discovery that a
small-fruited plant, obviously a kumquat, was masquerading under the name
Atalantia hindsii led Swingle to a study of the group, with the result
that in 1915 he proposed a new genus, Fortunella, to include all the
species of kumquats that had hitherto been placed in Citrus and
Atalantia. Important characters were brought to light that
separated Fortunella sharply from both Citrus and
Atalantia. Further studies have disclosed the fact that a
number of varieties of kumquats found in cultivation in China and Japan are
merely hybrids due to chance cross-pollinations by insects in village or
dooryard groves. The Meiwa variety (called Chintan in China), at
first considered by Swingle to be a good species (Fortunella
crassifolia), and the Changshou kumquat, named as a species (F.
obovata) by Tanaka, are both doubtless mere garden hybrids not entitled to
rank as species. This decision helps greatly in clarifying the
taxonomic status of the genus.
There are also
intergeneric hybrids resulting from cross-pollinations between Fortunella
and Citrus found in cultivation in China and neighboring
countries. One such hybrid, the Calamondin, has been erroneously
named as a species of Citrus (it was called C. microcarpa by Bunge
and C. mitis by Blanco). However, the whole complex
intergeneric hybrid problem of Fortunella has been materially clarified
by the making of many accurately safeguarded cross-pollinations of
Fortunella with Citrus, Poncirus, Microcitrus, and
Eremocitrus. This work, conducted under Swingle's direction
in the Agricultural Research Service, led to the creation of a bewildering
group of strange hybrids that would have been very disturbing to the taxonomist
if their origin were not definitely known. They are discussed below
under Fortunella hybrids.
Several
species of Fortunella have developed a degree of resistance to winter
cold and at the same time an even more important physiological peculiarity,
namely, a very pronounced winter dormancy that permits them to pass through
weeks of warm weather without starting growth or flowering. This
quality, possessed in much higher degree by the kumquats than by any other
citrus fruit tree (not excluding the winter-hardy Poncirus), makes them
of prime importance in the breeding of new types of hardy citrus fruit trees
(especially for producing acid citrus fruits) able to grow in much colder
regions than the lemon or lime (both notoriously deficient in winter dormancy
and hence easily pushed into growth by a few days of untimely warm weather in
winter or early spring).
Fortunella
hybrids will in all probability be produced in large numbers in the
future. Their study is likely to prove of great importance in
bringing about a just appreciation of the hybrid problem in the taxonomy as
well as in the breeding of citrus fruit trees. Since
Fortunella, with its few species and its already well-understood
intergeneric hybrids, constitutes a veritable microcosm of the citrus world, it
will doubtless prove of great help in understanding the much more complex and
still only imperfectly studied species and hybrid problems in the genus
Citrus itself. A key to the subgenera and species of Fortunella is
presented.
Subgenus Fortunella
Subgenus Eufortunella Swing. in Webber
& Batchelor, Citrus Indus. 1:346. 1943.
Ovary with 4-7 locules; fruits more than 1.5 cm
diam., with many pulp-vesicles.
1. Fortunella margarita (Lour.) Swing. Jour.
Wash. Acad. Sci. 5:170. 1915. Citrus margarita Lour. Fl. Cochinch.
2:467. 1790; C. aurantium [var.] olivaeformis Risso ex
Loisel.-Deslong. Nouv. Duhamel 7:95. 1816; C. aurantium var.
japonica Hook. f. Curt. Bot. Mag. 3 ser. 30:pl. 6128. 1874; C.
aurantium subsp. japonica var. globifera subvar.
margarita Engl. Die Nat. Pflanzenfam. 3(4):199. 1896.
Illus. Hooker f. loc. cit. pl. 6128 (col.); Siebold &
Zuccarini, Fl. Japon. 1:35, pl. 15, fig. 3. 1835; W. G. Smith, Gard.
Chron. 2 ser. 2:337, fig. 72. 1874; same figure, ibid. 3 ser.
7:393, fig. 58. 1890; also same figure, redrawn in Gartenflora
31:pl. 1097 (col.). 1882; Swingle, loc. cit. 5:171, fig. 2;
idem, in Bailey, Stand. Cycl. Hort. 2:1269, fig. 1563(1); 1270,
fig. 1564(1). 1915; Hume, Cult. Citrus Fruits 118, fig. 76.
1926.
Type.—Canton, China, cultivated
(coll. by Loureiro, but lost).
Distribution.—Known only in cultivation; doubtless native in southeastern
China.
Common name.—Oval kumquat.
Loureiro's original description (translated from
the Latin) reads as follows: "Differs from the species of Citrus in its
ascending, spiny branches, its linear petioles, its 5-locular, oblong
fruits.
"Habitat and notes: A tree 4 ft. [1.22
m] high, branches spiny, ascendant; leaves lanceolate, entire, shining, few;
petioles linear; flowers white with 5 petals, fragrant; peduncles sparse,
few-flowered; fruit oblong-ovate, yellowish-red, glabrous, 8 lines [16 mm]
long; 5-locular; peel thin, pulp vesicular, sweet, edible.
"Habitat: Canton, China, not rare.
Not seen by me in Cochin China."
Hume (1926)
gave the following description: "Tree dwarf, eight to twelve feet, bushy; young
branches somewhat angled, light green; leaves 1 1/4-3 1/2 X 3/8-1 1/4 inches
[3-8.8 X 1-3 cm], lanceolate, apex obtuse; base acute or obtuse; margins
crenate down about half-way from the apex; veins inconspicuous, surface dark
green, glossy; lower lighter; borne on rather stout, usually very slightly
margined, petioles, 1/4 to 5/8 inch [6-16 mm] in length. Fruit
small, obovate or oblong, 1 1/4-1 3/4 X 3/4-1 3/16 inches [3-4.4 X 1.8-2.5 cm],
golden yellow; stem short; calyx small; rind smooth, aromatic, spicy; oil
glands large; juice acid, sparse; sections usually five; seeds 2 to 5, oval 1/2
inch [13 mm] long, greenish; cotyledons two, green; season October-January.
The oval kumquat differs from the round kumquat
chiefly in the following respects: (1) the leaves are larger, more acute at the
base, less pallid, and more veiny below; (2) the ovary has usually four or five
(not four to seven) locules; (3) the fruit is oval, not globose; (4) the style
is persistent, not caducous; (5) the seeds are larger and especially longer,
with a rougher testa. It differs also in being distinctly more
vigorous and attaining a greater height (3 to 4 m); in the somewhat brighter
orange color of its fruits; and especially in the harsher, more biting flavor
of the peel, which evidently contains an ethereal oil more like that of the
common orange than is that of the round kumquat.
The type specimen of Citrus margarita
seems to have been lost, but Loureiro's description is very good and can apply
only to an oval kumquat very similar to the one commonly cultivated in all the
warmer parts of the world.
Robert Fortune, the
celebrated explorer of the tea regions of China, brought back plants of the
oval kumquat from China and delivered them to the Royal Horticultural Society's
Garden in London on May 6, 1846. He stated (1848) that he had
found numerous groves of kumquats thriving to perfection, along with tea, on the
lower hills of Chusan Island (off the coast of Chekiang Province, near latitude
30°) where the climate is far too cold to permit the culture of ordinary sweet
oranges or mandarins. He added: "this shows, therefore, that the
Kum-quat is of a much hardier nature than any of the plants belonging to the
orange tribe with which we are acquainted in gardens."
In a later Publication Fortune (1870, p. 50)
asserted that the kumquat needs a hot summer, 80° to 100° F and "will bear
without injury 10° to 15° of frost, and perhaps even a lower temperature."
The oval kumquat reached the United States by
1850 (see Hume, 1926, 114), but the round kumquat was not known in either
Europe or the United States until near the end of the nineteenth century.
2.
Fortunella japonica (Thunb.) Swing. Jour. Wash. Acad. Sci.
5:171. 1915. Citrus japonica Thunb. Nov. Act. Soc. Sci. Upsal.
3:199. 1780; also Fl. Japon. 292. 1784; C. madurensis Lour. 1790;
C. inermis Roxb. 1832; C. aurantium subsp. japonica var.
globifera Engl. Die Nat. Pflanzenfam. 3(4):199. 1896. Illus.
Thunberg, Icon. Pl. Japon. 2:pl. 5. 1800; Siebold & Zuccarini,
Fl. Japon. 1:pl. 15. 1835; Fortune, Jour. Roy. Hort. Soc. London
3:240. 1848; Swingle, loc. cit. 5:168, fig. 1; p. 171, fig.
3; idem, in Bailey, Stand. Cycl. Hort. 2:1269, fig. 1563(2); 1270,
fig. 1564(2). 1915; Hume, Cult. Citrus Fruits 116, fig. 74.
1926.
Type.—Japan (in culture)
(Thunberg). Herb. Thunberg, Bot. Mus.
Uppsala.
Distribution.—Known only in
culture, doubtless native in southern
China.
Common name.—Round kumquat.
The first fairly complete description of this
species, under the name Citrus japonica, was published in 1784 by
Thunberg in his Flora japonica; it reads in translation, as follows:
"petiole winged, leaves acute, shrubby stem. Japanese: Kin Kan,
vulgo Fime tats banna, Kaempf., Am. ex., Fase. V. p. 801. Growing
here and there, often cultivated for its little fruits. Stem
shrubby, compressed-subangulate, erect, glabrous, scarcely a foot
high. Branches and branchlets alternate, compressed-angular,
spinose, glabrous, erect, green. Spines solitary, in the axils of
the leaves, erect divergent. Leaves few, with winged petioles,
ovate, somewhat acute, entire, somewhat concave, glabrous, dark green above,
paler below, erect, spreading, 1 in. [2.5 cm] long, with very minute glands
('poris'). Petioles winged. 1 line [2 mm]
long. Flowers axillary, often solitary, rarely paired, pediculate,
nodding. Pedicels glabrous, hanging down, 1 line [2 mm]
long. Parianth, 1-phyllous, green, glabrous, minute,
5-toothed. Petals 5, white, oblong, somewhat concave, spreading,
somewhat claw-like. Filaments 19, subulate, compressed, erect, in 5
more or less coherent bundles, connate, forming a cylinder, shorter than the
corolla, nearly equal in length, white. Anthers oblong, small,
yellow. Ovary superior, subglobose, glabrous. Style
solitary, cylindrical, slightly shorter than the stamens, greenish
white. Stigma simple, globose, yellow, striate, many-locular
within. Fruit with fleshy peel, vesicular pulp, 9-locular
[sic], the size of a cherry. Differs from the other species
of Citrus especially in being a very small shrub with minute fruits; thus
it can scarcely be considered as a variety of orange. It resembles
Citrus medica in the axillary flowers, but differs in the winged
petioles; it differs from the orange in the axillary flowers, which are solitary
or paired, never in panicles. Fruit ripens in December and January,
is very sweet, agreeable and edible."
This
description was evidently drawn from a very small plant, perhaps an artificially
dwarfed one, such as are commonly grown in Japan. The very small
leaves with axillary spines, and the fruits "the size of a cherry" strongly
indicate that Thunberg was describing the round kumquat and not the oval
kumquat (F. margarita). His plate of this plant published in
1800 shows a flowering twig with small leaves and well-developed axillary
spines.
Hume (1903 and 1926) described this
species as follows:
"Tree similar to Nagami
[F. margarita], except that it is slightly thorny, and has the leaves
somewhat smaller and rounder at the apex. Leaves oval; apex obtuse;
margin crenate halfway down the length; veins slightly more conspicuous than in
Nagami; borne on short rigid, inconspicuously winged petioles, 1/4 to 1/2 in.
[6-13 mm] in length. Fruit spherical or somewhat oblate, 1 to 1 1/4
inches [2.5-3 cm] in diameter; golden yellow, short stalked; calyx small; rind
smooth, thin, spicy to the taste and aromatic when bruised; oil cells large;
pulp sparse; juice acid; sections four to seven; seeds one to three, small,
oval, greenish; cotyledons two, greenish. Season same as
Nagami."
SPECIES OF DOUBTFUL VALIDITY
3. Fortunella polyandra (Ridl.) Tan. Studia Citrol.
6:33. 1933. Atalantia polyandra Ridl. Fl. Malay Penin.
1:359. 1922; Fortunella swinglei Tan. Bul. Soc. Bot. France
75:714. 1928; Citrus polyandra (Ridl.) Burkill, Gard. Bul. Straits
Settl. 5:219. 1931; C. swinglei Burkill ex Harms, in Engler, Die
Nat. Pflanzenfam. 19a:459. 1931.
Type.—Malay Peninsula (Ridley). Herb. Bot. Gard.,
Singapore.
Distribution.—Malay
Peninsula; Southern China: Hainan Island.
Common name.—Malayan kumquat.
Ridley
described this species as follows, under the name Atalantia polyandra:
"Unarmed glabrous bush, branchlets smooth with a ridge decurrent from the
petiole; leaves thinly coriaceous, unifoliolate; leaflets lanceolate, bluntly
acuminate, base narrowed; main nerves slender, about 10 pairs; intermediate
nerves nearly as thick, numerous, irregular, all inarching; [leaflet] 4-6"
[10-15 cm] long 1.25-2.75" [3.2-7 cm] wide; petioles 0.25 to 0.75" [6-19 mm]
long, winged above; flowers few, usually 2 in an axil, pedicels 0.25" [6 mm]
long; calyx lobes 5, ovate-acute, short; petals 5, linear-oblong, blunt, 0.5"
[11 mm] long; stamens 24, connate, in a tube, some shorter than the others,
tube nearly 0.5" [11 mm] long, cylindric; anthers small-ovoid; disc fleshy,
cushion-shaped, forming a gynophore; ovary on the disc, oblong, glandular,
5-celled; style short, stout; stigma oblong, clubbed, 5-ribbed; fruit globose
with numerous large glands 1.75" [4.5 cm] through (not ripe)."
In his description of Fortunella swinglei,
based on a specimen from the Malay Peninsula, Tanaka stated: "fruits small,
globose, 1.5 cm diam., with 5 locules; peel thin, smooth, minutely punctate;
seeds ovoid, 10 X 15 mm, flattened, apex rounded, base very acute."
A kumquat collected in May, 1922, by F. A.
McClure (No. C.C.C. 9449) along a woodcutters' path in dense forest on the
south slope of Five Finger Mountain, now in the National Arboretum Herbarium at
Washington (C.P. Or. Herb. No. 7714), was identified by Tanaka (slip SW-316) as
Fortunella swinglei. The leaves are slender, lanceolate,
acuminate, with the very tip emarginate; the petioles are slender, 12-14 mm
long, and very narrowly winged above. Fruits are not
known. It looks like the wild form from which the Malayan kumquat
has been developed by culture or by hybridization.
This species is as yet inadequately
known. The cultivated form common in the Malay Peninsula, where it
is called limau pagar, or "hedge
lime," has much larger fruits than any other species of Fortunella,
and likewise a thinner peel. It may possibly prove to be a limequat,
i.e., a hybrid of a Fortunella and some variety of Citrus
aurantifolia, but, if so, it is abnormal in having only five locules in the
ovary and only two ovules in each locule. It also has an abnormally
high number of stamens, which may indicate hybridity.
Subgenus Protocitrus
Subgenus
Protocitrus Swing. Jour. Wash. Acad. Sci. 5:174. 1915.
This subgenus differs from the subgenus
Fortunella: (1) in having the ovary hypomerous (3- or 4-loculed, not 4-
to 7-loculed); (2) in having in mature fruits on the inside of the ovary wall
between the stalks of the pulp-vesicles a number of minute, wart-like, pale
yellow, cellular masses; (3) in having the dissepiments of the fruit dry, and
the peel thin and not very fleshy; (4) in having shorter, broader, more
brachytic flowers; (5) in having leaves with the veins more prominent on both
faces, and less pallid below.
The two most
important characters distinguishing the subgenus Protocitrus from the
subgenus Fortunella are the few-loculed ovary and the dimorphic emergenzen arising from the ovary wall of the fruit,
viz., ordinary pulp-vesicles and verruciform tufts of loosely aggregated, more
or less colored cells. These curious organs are analogous to the
short-stalked, clavate slime glands found in Citrus, but are much more easily
seen as they are not intermingled with the developing pulp-vesicles; they also
persist longer, being visible in half-grown fruits.
4.
Fortunella hindsii (Champ.) Swing. Jour. Wash. Acad. Sci.
5:175. 1915. Sclerostylis hindsii Champ. ex Benth. Hook. Jour.
Bot. 3:328. 1851; Atalantia hindsii (Champ.) Oliv. ex Benth. 1861.
Illus. fig. 3-32.
Type.—Hong
Kong (Hinds, no number, coll. 1841). Herb.
Kew.
Distribution.—Hong Kong; China:
Kwangtung and Chekiang provinces.
Common
name.—Hongkong wild kumquat.
A spiny shrub
or small tree; twigs slender, angled when young; leaves oval-elliptical,
tapering sharply at both ends, dark green above and faintly venose, paler and
not venose below; petioles winged, often emerging into the lamina of the leaf
without a separative joint; flowers short, broad, not opening very widely;
pistil very short; style shorter than the ovary; stigma large, cavernous; ovary
with 3 or 4 locules; ovules 2 in a locule; fruits small, 1-1.5 cm diam.,
subglobose, bright orange or scarlet-orange when ripe (the color of a tangerine
orange); pulp-vesicles very few, small, fusiform; seeds thick, oval or ovate in
outline, plump, 9-11 X 7-8 X 5-6 mm, pistache green in section.
The Hongkong wild kumquat, Fortunella
hindsii, differs from the round kumquat (F. japonica) and the oval
kumquat (F. margarita) in a number of morphological characters, some of
them being of decided taxonomic significance in this group. Longley
(1925, p. 347) found the pollen mother cells of this species to contain eighteen
instead of nine chromosomes as in all other wild-growing species of the orange
subfamily. This plant is in the tetraploid state, very probably in
an autotetraploid state.
Frost (1926, pp.
380-81) found that tetraploid forms of the commonly cultivated species of
Citrus (sweet orange, tangerine and mandarin, grapefruit and lemon) show
"unfavorable tree and fruit characters," such as thickened leaves, lower vigor,
slowness to bloom, coarser rind, and lowered acid content in the fruits.
Although F. hindsii has long been known
to the Chinese and is not uncommonly cultivated in both China and Japan, it was
not discovered by Europeans until about a century ago. At first it
was not recognized by taxonomic botanists as being closely related to the other
species of kumquats (then placed in Citrus) but was considered to be a
species of Atalantia (probably because of the small size of the fruit),
in spite of the fact that the stamens are four times as numerous as the petals,
whereas in Atalantia the stamens are only twice as numerous as the
petals.
The earliest known Chinese reference
to Fortunella hindsii is found in the first monograph on citrus fruits
ever published in any country, the famous Chü lu (Orange Monograph) of
Han Yen-chih (1923), first published in 1178. In the translation
made by Hagerty, the paragraph about wild kumquats reads as follows: "Chin
chü [golden chü orange]. The chin chü grows along
the sides of mountain paths; compared with the chin kan [golden mandarin,
the ordinary round kumquat], this fruit is very much smaller, but is similar in
color and form…We sometimes find the tree bearing enough to fill several pint
measures. The pulp segments of the chin chü cannot be
divided. It has but one seed but the taste of its pulp is such that
it cannot be eaten. The chin chü trees are only suitable to
be placed in pots on balcony railings. Many gardeners cultivate and
sell them for this purpose."
Han Yen-chih
then went on to say that this little tree is also called shan chin kan
(mountain golden mandarin), meaning wild kumquat, and quoted a song about it
written by Chou Mei-ch'êng, a poet of the Sung dynasty, who must have written
before A.D. 1178, the year Han Yen-chih’s work was published. A few
centuries later, in the Ming dynasty (1368-1644 A.D.), we find Chinese authors
using the name chin tou (golden bean) for this species. This
name is found in Min shu (Records of Fukien), compiled by Ho Chiao-yüan,
who lived toward the end of the fifteenth century. In describing
the kinds of kumquats, he wrote: "there is also the chin tou…growing in
the mountain forests; when preserved in honey they are very good."
T’u Pên-tsun (1600?), an author of the Ming
dynasty, in an essay, P'ing shih yüeh chih (Calendar of Garden Flowers),
named clearly three kinds of kumquats: "the three little friends, the chin
tou [or golden bean, Fortunella hindsii], the chin kan [or
golden mandarin, F. japonica] and the chin chü [or golden orange,
probably F. margarita]." Swingle (1922 and 1929a)
pointed out that this was apparently the first definite recognition of the fact
that these are different kinds of kumquats, now considered to be three distinct
species.
4a. Fortunella hindsii var.
chintou Swing. Jour. Arnold Arbor. 21:130. 1940. Illus.
fig.
3-33.
Type.—Plant introduced from
Japan and grown in the U.S. Plant Introduction Greenhouse near Glen Dale,
Maryland (C.P.B. 907, F.P.I. No. 71241). Herb. Natl. Arbor.,
Washington, D.C.
Distribution.—Cultivated in the warmer regions of China and Japan as an
ornamental plant. Doubtless native, or originated under culture in
southeastern China.
Common
name.—Golden-bean kumquat.
Differs from
the parent species, F. hindsii, in having larger, thinner, somewhat
narrower leaves, 3.5-8 X 1.5-2.5 cm, shorter, more slender spines, and larger,
slightly depressed, globose fruits, 12-15 mm in diam., and in having the normal
diploid number of chromosomes (9 in the gametes and 18 in the somatic cells)
instead of twice as many (18 and 36), as the parent species has.
It has also distinctly smaller flowers with petals 5-6 X 2.5-4 mm instead of 6-7
X 4-5 mm, as in the tetraploid species growing alongside; blunter, much shorter
calyx lobes (0.5-0.6 mm long instead of 0.8-1.2 mm, as in the species) and a
somewhat narrower and evidently shorter disk than in the parent species.
This striking dwarf kumquat is probably a
cultivated variety derived from the wild species. It is unique
among the Citrus Fruit Trees in being the diploid
state of a normally tetraploid wild species. As is remarked
under Citrus, autotetraploid cytonomic states of the commonly cultivated
species of Citrus have been found by Frost (1925, 1926, 1938a,
1938b) to occur not uncommonly in California among seedlings of the sweet
orange, tangerine, mandarin, grapefruit, and lemon. Such
autotetraploid seedlings usually show many or all of the following
characteristics, as compared with the parent species: a tendency to slower
growth, later bearing, and being less floriferous and less fruitful; also
thicker and broader leaves and winged petioles, more thorny twigs, shorter
fruits having thicker rind with larger oil glands, less juice, and larger,
especially longer seeds.
Swingle was able to
observe minutely this cultivated diploid variety during a trip to Japan and
China in 1926 and to bring back living plants with him to this
country. This cultivated form, called by the Chinese
Chin-tou (pronounced kindzu in Japan), meaning "golden bean," has
just such characters as would be expected in a diploid reversion from an
autotetraploid species. It may, however, prove to be a cultivated
form of a species which still persists as a diploid plant.
THE THREE CLASSES OF FORTUNELLA
HYBRIDS
The hybrids of Fortunella are many and are
both intrageneric (inside the genus) and intergeneric (between Fortunella
and other genera). The hybrid question comes up at every turn
in classifying cultivated citrus fruit trees and is the chief hindrance to
phylogenetic taxonomy of the species of Citrus and
Fortunella. It can be given a simpler and at the same time a
more comprehensive treatment in a discussion of Fortunella than of
Citrus, for the latter has many more species and many more hybrids that
are often less easily recognized.
There are at
least three distinct classes of Fortunella hybrids. (1)
intrageneric hybrids between species of Fortunella; (2) bigeneric hybrids
between species of Fortunella and species of Citrus,
Poncirus, Microcitrus, or Eremocitrus; and (3) trigeneric
hybrids.
INTRAGENERIC HYBRIDS
Two cultivated
varieties of kumquats, the Meiwa (or Chintan) kumquat of China and Japan, widely
distributed, and the Changshou (longevity) kumquat growing in Chekiang and
Fukien provinces of China, are probably to be classed as hybrids between two
species of Fortunella.
Meiwa kumquat.
According to
Tanaka (1922, p. 248), the Meiwa kumquat was so named in Japan because it was
introduced from China during the reign of the Japanese emperor Meiwa
(1764-1771). It is usually called Chintan (golden bullet) in China
and is widely grown in Chekiang Province (Hu, 1934, pp. 22-23).
The Meiwa kumquat is figured and described in
Swingle (1915c, pp. 172-74, fig. 4) and in Hume (1926, p. 117, fig.
117). The fruits are broadly oval or sometimes subglobose, 25-35 X
25-28 mm, ordinarily with 7 segments, and usually some of them
seedless. The peel is very thick, nearly twice as thick as that of
either the round or the oval kumquat, and is very sweet and good
flavored. The leaves are also thicker than those of the common
round or oval kumquats and have 4-5 or even more layers of palisade cells on
the upper surface instead of 2 or 3 layers.
The nearly thornless twigs, the often nearly seedless fruits and the increased
thickness of the peel, the irregularly thickened segment walls, and the
thickened leaves with more layers of palisade cells are in a way an
intensification of kumquat characters that may have resulted from a chance
hybridization of the round and the oval kumquat, or a back-cross with a
Fortunella of a Citrus-Fortunella hybrid. The broadly
oval shape of the fruits of the Meiwa kumquat is about midway between the oblong
fruits of Fortunella margarita and the globose fruits of F.
japonica.
Swingle named this variety as a
new species, Fortunella crassifolia (1915c, p. 172), but suggested
that it might be of hybrid origin. He later considered it to be a
garden hybrid not entitled to rank as a
species.
Changshou kumquat.
The
Changshou (longevity) kumquat is a dwarf variety commonly grown as a potted
plant in China, in the provinces of Wenchow, Fuchow, Chekiang, and
Fukien. A condensation of Hu's description (1934, p. 24)
follows.
Fruits obovate with the tip concave,
about 3 cm long and 3 cm wide, with 8 segments. The peel is very
thin (1.5 mm) and has the same flavor and odor as that of the round kumquat,
F. japonica. The seeds are few (rarely more than 2 or 3) and
polyembryonic. This variety was fully described by Tanaka
(1932c, pp. 151-52, in Japanese; and 1933a, pp. 38-40, in
English). He reported the plant as thornless, the fruits as "about 3
cm broad, and slightly over 3 cm in height, obovoid, sometimes nearly globose,
but broadest at the apical part; apex abruptly and rather deeply
concave…" The peel is said to be 2.5-3 mm thick, soft, sweet and
edible; segments 5-6; the pulp, "juicy, subacid, refreshing." Seeds
sometimes absent but often rather abundant if present, polyembryonic with more
embryos than those of the Meiwa kumquat.
The
dwarf habit, absence of spines, the frequent seedlessness of the fruits, the
concavity at the apex, and the large number of embryos in the seeds, all point
to this as being a chance hybrid between two species of
Fortunella. It was named as a species, F. obovata, by
Tanaka (1933a, p. 38), but specific names for forms suspected of being
garden hybrids or mutations are of doubtful taxonomic validity and should not
be accepted if proof is lacking that the plants are not mere garden hybrids,
propagated only by man's help.
It is probable
that other kumquat hybrids exist in China, since differences in the articulation
of the petioles with the leaf blades are observable in herbarium specimens of
Fortunella collected in China.
BIGENERIC HYBRIDS OF FORTUNELLA WITH CITRUS AND PONCIRUS
The citrus experts of the Agricultural Research
Service, using adequate precautions to exclude all stray pollen, have made a
number of bigeneric hybrids of Fortunella. Apparently, some
such hybrids have arisen without human help in the dooryard or village groves
of mixed genera, species, and varieties of citrus fruit trees. It
is probable that some of these chance seedlings found in the Far East are
second-generation kumquat-citrus hybrids or else backcrosses of an F1 Fortunella X Citrus hybrid on one of the
parental species.
Limequats [Citrus aurantifolia X Fortunella
sp.].
These hybrids have been described by
Swingle (1914-1917, vol. 4, p. 1882, fig. 2176); by Swingle and Robinson (1923,
pp. 235-38, pls. 4, 5); by Hume (1926, pp. 134-35, fig. 88); and by Hodgson (see
chap. 4, this work).
Most of the named
varieties of limequats, Eustis, Lakeland, etc., are hybrids of Fortunella
japonica with Citrus aurantifolia 'Mexican,' the round kumquat being
the pollen parent. Another named variety of limequat, the Tavares,
has the oval kumquat as the pollen parent (Citrus aurantifolia 'Mexican'
X Fortunella margarita).
Orangequats [Citrus reticulata c. satsuma X
(Fortunella japonica X F. margarita 'Meiwa’)].
The orangequat is a complex, artificial,
safeguarded hybrid between the satsuma orange and the Meiwa kumquat (the
latter, the pollen parent, is here considered a hybrid of the round and the oval
kumquat). A named variety of this hybrid, the Nippon orangequat,
was described in Robinson and Savage (1934, p. 11). The fruit is
broadly oval or obovate in shape, 3.8 to 5.0 cm in diameter by 5 to 6.3 cm in
length, with thick, spongy peel of mild flavor and with acid
pulp.
Citrumquats [Poncirus trifoliata X Fortunella
japonica].
The citrumquat proved an
exceedingly difficult hybrid to make. For several years no viable
seeds were obtained. When at last a few were secured, the young
seedlings all died promptly. Finally, in 1909, Swingle found a few
belated "June blooms" on a Poncirus trifoliata tree, nearly all of which
set fruit when pollinated with pollen from Fortunella japonica.
From these cross-pollinated trifoliate oranges nearly a hundred seeds
germinated. Many of the seedlings died while still very young but
a few lived and a very few made healthy, vigorous plants that were obviously
hybrid plants and not nucellar seedlings of the female parent. (See
fig.
3-34.)
This hybrid looks much like a
citrange in twigs and foliage.
The citrumquat
is a combination of two citrus fruit trees possessing to the highest degree two
very different kinds of hardiness. Poncirus trifoliata is a
deciduous tree able to endure winter temperatures as low as -23° C or -25° C
(-10° F to -13° F), whereas the kumquat, with its highly developed winter
dormancy, is able to withstand long spells of warm weather in winter or early
spring without putting out tender new growth easily killed by temperatures of
even only a few degrees below the freezing point. This winter
dormancy of the kumquat was noted by Swingle and Robinson (1923, p. 230) in
their discussion of citrangequats.
The
extreme difficulty experienced in getting healthy hybrids between Fortunella
and Poncirus is further proof of the remote degree of relationship of
these genera. However, failure to get viable hybrids on the first
trial should not be taken as conclusive proof that such hybrids cannot be
obtained if the parent plants (particularly the seed parent) are in proper
condition when cross-pollinated.
Calamondin (orangequat?) [Citrus reticulata var.
austera ? X Fortunella sp. ?].
This Chinese citrus fruit tree, illustrated in figure 3-35, widely cultivated in the Philippines and also grown
in Hawaii and the United States, is very probably an orangequat that arose in
China by insect cross-pollination of a sour, loose-skinned mandarin orange and
a kumquat, perhaps Fortunella margarita. In the Philippines,
it is known under the name Calamonding. This hybrid was described
under the name Citrus mitis Blanco by Swingle (1914-1917, vol. 2, p. 784)
and by Hume (1926, pp. 133-34, fig. 87) and under the name Citrus
microcarpa Bunge by Tanaka (1933b, p. 184). It was
figured in Ochse (1931, pp. 131-32, col. pl. 50). It is reasonably
certain that the Calamondin is a Citrus X Fortunella hybrid and
should not be considered as a valid species (see Swingle, 1942, p. 26).
This hybrid has depressed-globose fruits with a
very thin peel, that becomes loose as the fruit ripens, and with intensely acid
pulp. The segments number only seven to ten. It may
possibly turn out to be a back-cross of an F1
Citrus X Fortunella hybrid on Citrus reticulata.
This could probably be determined by experimental hybridization.
This hybrid enters into the parentage of the interesting trigeneric hybrid, the
Altamaha or Glen citrangedin.
Tetraploid
Calamondin.
This interesting form of the
Calamondin was discovered in Formosa by N. Nakamura, one of Tanaka's graduate
students. He published (1934) an account of his examination of a
large number of cultivated forms of Citrus, including at the end of the
list several forms of the Calamondin and two other hybrids. In all,
he examined seventy-six species or cultivated varieties of Citrus and
four citrus hybrids. Only one of these varieties (the one now in
question) showed the tetraploid number of chromosomes. He called
this variety "Shikinari-mikan" and considered it to be a tetraploid form of the
Calamondin. He noted that the pollen mother cells show 18 bivalent
chromosomes in the first division of the pollen mother cell, and 18 monovalent
ones in the second division (Nakamura, 1934, p. 169, figs. 9, 10).
He stated that the characters of this variety correspond in general to those of
the diploid form of Calamondin, commonly cultivated in Japan; however, the
tetraploid variety has "fruits…larger in size and sweeter, and…leaves…much
thicker than those of the diploid individuals.…Its pollen grains are also
larger than those of the diploid," but more of them (about 30 per cent) are
sterile.
Unlike many tetraploid forms of
Citrus, this tetraploid Calamondin grows as vigorously as the diploid
form, or even more vigorously.
COMPLEX BIGENERIC HYBRIDS
Procimequat [(Citrus aurantifolia 'Mexican’ X Fortunella
japonica) X Fortunella hindsii].
This very interesting complex hybrid, which has a triploid chromosome number,
is the result of a carefully safeguarded cross-pollination of the Eustis limequat with Fortunella hindsii, a tetraploid species
made by Eugene May and the writer expressly to obtain a triploid
hybrid. Longley (1926, pp. 543-45, fig. 1) found it to be triploid,
with 27 chromosomes in the somatic cells (18 supplied by the male parent, the
Hongkong wild kumquat, and nine by the limequat).
The limequat fruits have from six to nine
segments, as might be expected from a hybrid of the round kumquat (with four to
seven segments) with the Mexican lime (with 10 to 12 segments). The
Hongkong wild kumquat fruits have only three or four segments. The
ovaries of the procimequat hybrid under consideration usually show from four to
five segments.
The leaves of these hybrids are
small but some of them show fairly vigorous growth (see fig. 3-36). The fruits set abundantly even on small
young plants and are small and subglobose, much like those of Fortunella
hindsii but a little larger and a much paler orange in color when
ripe. These fruits are not seedless, as was expected, but produce
some nucellar bud embryos, as do many citranges after the development of the
ovules has been stimulated by pollination. Triploid limes are
usually seedless.
This hybrid is interesting
because it throws light on bigeneric Fortunella X Citrus
back-crosses such as are possibly represented by the Malayan hedge lime discussed. The procimequat is in
reality intermediate between a true bigeneric back-cross and a trigeneric
hybrid, because Fortunella hindsii belongs to a subgenus,
Protocitrus, with many important taxonomic characters separating it from
the true Fortunella species placed in the subgenus Fortunella.
The name "procimequat" (given here for
convenience) is derived from Pro[to]c[itrus X
L]imequat.
TRIGENERIC HYBRIDS OF FORTUNELLA
Several remarkable trigeneric hybrids having Fortunella as one parent
have been made by the citrus experts of the former Bureau of Plant
Industry. These hybrids are of great interest because of the light
they throw on the origin of strange complex hybrids that arise without human
aid in the dooryard citrus groves of China, Japan, and
India.
Citrangequats [Fortunella sp. X (Citrus sinensis
X Poncirus trifoliata)].
These were
the first trigeneric hybrids to be produced artificially from definitely known
parents. Swingle succeeded in making this cross in 1909 by using
pollen from Willits and Rusk citranges (Poncirus trifoliata X Citrus
sinensis) on properly safeguarded flowers of the oval kumquat (Fortunella
margarita) and the round kumquat (F. japonica). Several
of these citrangequats were described and illustrated by Swingle and Robinson
(1923, pp. 230-33, pls. 1, 2, 3). The Thomasville, Sinton, and
Telfair citrangequats, described and figured by Hume (1926, pp. 40-42, fig. 45),
are the best known of these hybrids.
Citrangequats vary greatly in size, color, flavor, etc. The
Thomasville citrangequat has obovoid fruits borne on long pedicels, 1.5 to 3 cm
long and 2 to 2.5 mm in diameter at the base, but swollen and pulvinoid at the
top, 8 to 10 mm, and permanently curving as the fruit matures until it hangs
down, making an angle of 45° to 90° with the base of the pedicel.
This character is unknown in any of the three parent species and shows how
taxonomically new characters can arise in complex hybrids (see further
discussion under Poncirus hybrids).
Citrangedins [(Fortunella sp. ? X Citrus reticulata
var. austera ?) X (Citrus sinensis X Poncirus
trifoliata)].
This highly complex
trigeneric and 4-specific hybrid is the result of a cross of the Calamondin,
itself a hybrid of Fortunella and Citrus, with the Willits citrange, a hybrid of Poncirus and
Citrus. It bears small, subglobose fruits of a brilliant
orange-yellow color. The fruit was described and figured by
Swingle, Robinson, and Savage (1931, pp. 14-15, pl. 9) and the plant’s extreme
resistance to cold was noted in Swingle, Robinson, and Savage (1932, p.
7). The 4-specific hybrid often shows one- or two-lobed leaves on
young vigorous shoots, and sometimes difoliolate or trifoliolate
leaves. The leaves on fruiting twigs are almost always unifoliolate
and look very like those of a Fortunella-Citrus hybrid. This
hybrid shows clear traces of all three genera that enter into its parentage but
is strikingly different from any species of Fortunella, Poncirus,
or Citrus.
If its parentage were not
known, the Altamaha or Glen citrangedin might even be considered a new genus,
as well as a new species, by those who name "chance seedlings" of obvious
hybrid ancestry as good species!
Faustrimedins [Microcitrus australasica X
(Fortunella sp. X Citrus reticulata ‘Calamondin’)].
The faustrimedin is a hybrid of the Australian
finger-lime with the Calamondin, itself a hybrid between the kumquat and some
variety of orange of the mandarin group (Citrus reticulata).
It is thus a trigeneric hybrid of Citrus, Fortunella and
Microcitrus. The faustrimedin has small leaves much like those
of the Australian finger-lime and is hardy like the Calamondin.
This hybrid, first made by Oliver (1911, p. 40,
pl. 1, fig. 2), may be of use in obtaining still more complex hybrids
containing the blood of Eremocitrus or Poncirus. It is
a vigorous grower and shows some promise as a rootstock. It is
easily propagated from cuttings. The fruits are like finger-limes
but are shorter, nearly seedless, and pleasantly acid.
XXII. Eremocitrus
Swingle
XXII. Eremocitrus
Swingle, Jour. Agr. Res. 2:86. 1914.
Type species.—Triphasia glauca Lindl. = Eremocitrus glauca
(Lindl.) Swing.
Distribution.—Australia: central and southern Queensland; northern New
South Wales. From near the coast at Broad Sound, Queensland, as far
west in scattered colonies as Long. 142° E. in New South Wales, or possibly
Long. 141° E. in Queensland.
Common
name.—Australian desert lime.
Leaves
gray-green, thick and leathery with appressed grayish hairs on both surfaces;
spines solitary in the axils of the leaves, sometimes lacking on older
branches; flowers single or few in the axils of the leaves, 3-5-merous, petals
narrowed at the base, stamens free, 4 times the number of the petals, ovary
with 3-5 locules, 2 ovules in each locule, style short; fruit oval or pyriform
with a fleshy peel (as in Citrus), having oil glands, pulp acid, with
stalked subglobose pulp-vesicles, seeds small with hard wrinkled testa;
cotyledons hypogeous, first leaves cataphylls.
This genus is in many ways the best characterized and most distinct of any of
the near relatives of Citrus. It is the only pronounced
xerophytic plant in the whole orange subfamily. Both leaves and
twigs are gray-green, like sagebrush; the leaves are small, thick, bifacial
with palisade parenchyma on both faces, and covered with a thick
cuticle. The stomata are deeply sunken in pit-like depressions in
the surfaces of both leaves and young twigs. There is a somewhat
persistent sparse coating of thick-walled appressed hairs on the leaves and
twigs not present in any other citrus fruit tree. In the event of
severe drouth, the leaves fall and the gray-green twigs carry on the very
restricted photosynthesis possible under such conditions. The young
plants have greatly reduced linear leaves that are, in fact,
cataphylls. The young seedlings grow very slowly until an extensive
root system is formed; then, if adequate moisture is found, the stem grows
upward as rapidly as that of any citrus tree and little by little larger leaves
are formed until some may measure 50 to 65 by 10 to 20 mm and nearly 2 mm
thick! The young plants have single sharp, stiff spines in the
axils of the leaves. These are usually 2 to 3 cm long.
On certain particularly vigorous wild trees growing in central New South
Wales, spines of enormous length have been produced. They are very
stout, somewhat up-curved, and reach a length of 6 to 9 cm with a vertical
diameter at the base of 5 to 7 mm and a transverse diameter of 3 to 4
mm. The flowers are much like those of Microcitrus but
smaller, usually 3- to 5-merous with six to ten entirely free stamens with
slender filaments. The ovary has three to five locules with two
ovules in each locale, a character not found in any other True Citrus Fruit
Tree except Fortunella! The fruit is small, subglobose,
ovoid or obovoid, 15 to 25 by 12 to 15 mm. The subglobose
pulp-vesicles have short, slender stalks and are loosely aggregated, not
cohering as tightly to each other as in the common citrus fruits.
The seeds are small and contain only a single embryo.
This remarkable xerophytic citrus fruit tree
grows wild in central and northern New South Wales and in southeastern
Queensland. In New South Wales, it is sometimes found in semiarid
regions even as far west as the Mootwingee Range (Lat. 31° 45' S., Long. 142°
10' E.), about 80 miles northeast of Broken Hill in extreme western New South
Wales.
Eremocitrus glauca (Lindl.)
Swing. Jour. Agr. Res. 2:88. 1914. Triphasia glauca Lindl. in
Mitchell, Jour. Exped. Trop. Austr. 353. 1848; Atalantia glauca (Lindl.)
Benth. Fl. Austr. 1:370. 1870. Illus. Fairchild, U.S. Dept. Agr.
Yearbook 1911:pl. 45, fig. 1. 1912; Swingle, loc. cit. text figs.
1-7; idem, in Engler, Die Nat. Pflanzenfam. 19a:332, fig.
153,a-o. 1931; Maiden, Forest Fl. N. S. Wales 7:245, pl. 248,
figs. A-H. 1921; Flanders, Calif. Citrog. 17:278, 6 unnumbered
half-tone text pls. 1932; fig. 3-37 this work.
Type.—Australia, Queensland, Dublin County, near junction of Maranoa and
Merivale rivers (Mitchell, 1846). Kew, Brit. Mus., and Gray Herb.
(Swingle, 1914b, p. 90).
Distribution.—Australia: central and southern Queensland; northern New
South Wales. From the Burdekin River (Lat. 20° S.) in northeastern
Queensland to the coast at Broad Sound (Lat. 23° S.), thence south along Long.
152° E. to the New England district in northeastern New South Wales, thence
southwest to Dubbo in central New South Wales (Lat. 32° 20' S., Long. 149° 35'
E.), thence westward (rather common) to Long. 147° E., or even 146° E., and
also occurring in scattered groups much farther west in the hills in the
semiarid region as far as Long. 142° E. in New South Wales and possibly Long.
142° E. or even 141° E. in Queensland.
Common name.—Australian desert lime.
The
typical form of this species as it occurs in southeastern Queensland,
Australia, is a small tree or a large shrub, sometimes growing as a thorny bush
only a few feet in height. F. M. Bailey described it as "a rigid
glaucous shrub of 2 or 3 ft., often armed with straight or incurved axillary
spurs of 1/2 in. or under…" However, Flanders (1932, p. 278)
figured a giant tree at Chinchilla, Layton County, in Queensland, some 130 miles
west-northwest of Brisbane, that had a single trunk, about 25 feet in height
and measuring 20 inches in diameter at 2 feet above the ground. The
round crown was about 28 feet across. Such a tree undoubtedly grew
where its roots had access to ground water.
The leaves are gray-green, very thick and leathery, usually oblong, linear- or
elongate-obcuneate, obtuse at the apex, often emarginate, 25-40 X 4-10
mm. Both surfaces show appressed, grayish hairs and have a thick
cuticle with sunken stomata. The leaves are more or less
paraheliotropic (on the edge to the light) and have no clearly defined upper and
lower surfaces (Swingle, 1914b, figs. 5, 6, 7). Flowers very
small, 3-5 mm long and 5-8 or 10 mm across, borne singly or in groups of 2 or 3
in the axils of the leaves on slender pedicels, 4-6 mm long, flowers
3-5-merous, with 4 (sometimes 3) times as many stamens as petals, stamens
short, filaments slender, 4-5 mm long (sometimes slightly united at the base
according to Bailey); pistil short, 3-4 mm long, ovary ovate with 3-5 locules;
fruits subglobose or obovoid, 7-12 X 8-10 mm, pulp-vesicles small, subglobose,
on short, slender stalks and nearly free from one another; seeds small, oval,
5-6 X 3-4 X 2.5-3 mm, testa rough, monoembryonic. Young plants
growing in the greenhouse at Washington, D.C., had very slender, straight or
slightly upcurved thorns, 25-35 mm long but only 1-1.5 mm diam.; young trees
growing out of doors at Riverside and Indio, California, show much stouter
spines, 2-3 mm diam., but seldom as long as 35 mm. The twigs on
young plants are usually very slender, 1-2 mm diam., and for several years
bear very narrow, slender leaves or cataphylls, 18-30 X 1-2 mm, and make very
slow growth; as soon as an extensive root system has developed, the young tree
grows upward rapidly, producing larger leaves on slightly stouter twigs, 2-3 mm
diam., that often show only very short spines or none.
The trees growing at Indio under irrigation
began to flower and set fruit when they were 12-15 feet high and about 8-10
years old. At that age the branches were comparatively short and
the trees compact with but little spread (at 20 years of age one tree was 20-22
feet high and the top spread 12-16 feet). The flowers appear very
abundantly in March, and by late May or early June the fruits begin to drop
while still green or slightly yellowish-green. The fruits are
juicy, thin-skinned, and rather pleasantly acid. Two forms occur:
one strain has subglobose fruits, the other obovoid or short-ovoid fruits.
The Australian desert lime, Eremocitrus
glauca, is a pronounced xerophyte, the only one found thus far in the orange
subfamily (fig. 3-38). It is able to withstand severe drouth and
hot dry winds. Under such conditions the leaves fall off and the
leafless gray-green twigs (resembling those of the smoke tree, Dalea
spinosa, and the paloverdes, common in the arid regions of the southwestern
United States and northern Mexico) carry on photosynthesis on a reduced
scale. The seedling develops an enormous root system before making
vigorous aerial growth and developing full-sized leaves (young seedlings bear
only very slender cataphylls). The roots
are able to endure rather high concentrations of salts in the soil moisture
and are much less susceptible to boron poisoning than are those of
Citrus. When in a dormant condition this species is able to
withstand, without injury, temperatures of ten or more degrees below zero
Fahrenheit (-24° C or lower). It probably possesses a higher zero
point of growth and, in consequence, a greater dormancy in late winter and
early spring than Citrus. It apparently ranks next to, but
does not equal, the kumquats in possessing both winter dormancy and resistance
to cold.
There is good reason to believe that
Eremocitrus has spread slowly westward in northeastern Australia over a long
period of time, probably some 20 million years or more, during which time this
plant has become adapted to withstand a semiarid climate and to grow in soils
having an appreciable saline content in the soil moisture. Trees of
Eremocitrus glauca have been found growing wild in northeastern Australia
up to east longitude 142° 10’, a distance inland of 800 to 960 kilometers (500
to 600 miles). The western half of this range falls in a region
classed as "Dry Steppe" by the Australian experts Andrews and Maze (1933),
whose maps show that Eremocitrus grows wild as far west as the climatic
zone which according to their designation has an average of from six to eight
arid months. Their No. 8 line (indicating eight arid months in the
year), which separates the steppe climate from the desert climate in Australia,
runs through the westernmost limit of the range of
Eremocitrus. Convincing proof of the drought resistance of
Eremocitrus is found in certain thrifty groves of the desert lime growing
in regions in Australia where salt bushes thrive and where trees of some other
species die during prolonged drought before the Eremocitrus trees show
even slight injury.16
In spite of years of effort on the part of
several generous Australian cooperators, the attempt to obtain seed from the
Eremocitrus trees growing wild in the drier parts of Queensland and New
South Wales proved unsuccessful up to 1941. A few mature fruits
were found west of longitude 148°, but they contained no viable
seeds. Fortunately, in December, 1941, C. T. White collected ripe
fruits of Eremocitrus glauca containing viable seeds in the vicinity of
Charleville, Queensland, in the Warrego River Valley (Long. 147° 40' to 145°
30' E.). The trees from which the seeds were obtained were growing
under semiarid conditions. These seeds germinated and the seedlings
grew well in the greenhouses of the U.S. Department of Agriculture near
Washington, D.C. The complete absence of seed in fruit of this
species growing in its natural habitat is apparently the rule, according to
information supplied in a letter dated December 2, 1964 to Dr. J. R. Furr, U.S.
Date and Citrus Station, Indio, California, from E. C. Levitt, New South Wales
Department of Agriculture, Sidney, Australia. As recently as 1964,
the E. glauca trees growing at the U.S. Date and Citrus Station have
been the source of seed supplied to Australian citrus specialists for their
studies on the suitability of E. glauca as a rootstock and its tolerance
to salinity.
Eremocitrus glauca can
serve as a rootstock for Citrus aurantium and other species of
Citrus; when used for the sour orange excellent growth was obtained in
the Washington greenhouses. Eremocitrus also grafts readily
on Citrus.
BIGENERIC AND TRIGENERIC HYBRIDS
OF EREMOCITRUS
Eremocitrus
trees growing near a collection of Citrus species and hybrids in the
U.S. Date and Citrus Station at Indio, California, have fruited heavily (fig.
3-39). A small percentage of the seedlings from them have proved
to be hybrids. In all, several score of such hybrids have been
found and many of them propagated from cuttings (fig. 3-40). Efforts to pollinate castrated and bagged
Eremocitrus flowers from Citrus or other genera have not so far
succeeded, probably because of the great profusion of blossoms produced by
Eremocitrus, of which doubtless only one in a hundred or so set
fruit.
Swingle believed that two kinds of
Eremocitrus hybrids are easily distinguished: bigeneric hybrids, in which
a Citrus species is the pollen parent, and trigeneric hybrids, which have
a citrange for the pollen parent.
In the
revision of this chapter, the junior author has maintained Swingle's
classification of bigeneric and trigeneric hybrids. However, it should be noted that
Dr. J. R. Furr, U.S. Date and Citrus Station, Indio, California, has
reservations concerning such classification. In a letter to Dr.
Walter Reuther, the editor of this volume, dated June 7, 1965, he stated:
I have some reservations regarding Swingle's
supposed eremolemon, eremorange, eremoradias, etc. As well as I can
remember the seedlings of the old E. glauca tree that used to grow at the
U.S. Date and Citrus Station looked about the same after the Meyer lemon tree
growing very close to it was removed as they did before its removal; but there
were grapefruit trees nearby all of the time, so it is possible that we were
getting E. glauca X grapefruit hybrids.
I have been puzzled by the E. glauca seedlings I have grown.
Usually there is a mixture of types in the open-pollinated seedlings grown from
our E. glauca trees ranging from vigorous, upright plants with lanceolate
leaves to very small, weak plants with thorny branches and only vestigial
leaves. The weak ones usually die in the flats before they are
large enough to plant in the field. I supposed that the weak thorny
plants were possibly selfed E. glauca seedlings and that the vigorous
ones were probably hybrids with citrus. We have never caged a tree
of E. glauca to see if it will produce any seed without
cross-pollination. One year I did pollinate flowers of E. glauca
with several different kinds of citrus. On different branches of
E. glauca, flowers were pollinated by hand with pollen from sweet orange,
pummelo, rough lemon, and Shekwasha. No seed [sic] were
obtained from flowers pollinated by pummelo. Seedlings that appear
to be hybrids were obtained from the other crosses; that is, the seedlings were
highly variable and there seemed to be family differences between the different
lots of seedlings of supposedly different pollen parents. In these
supposed crosses, the E. glauca flowers were not emasculated; the pollen
was just daubed on the stigmas of many open flowers, which may have been
self-pollinated at the same time.
I think
Swingle was probably right in supposing that many of the E. glauca
seedlings grown from the original old E. glauca specimen at the U.S. Date
and Citrus Station were hybrids. I doubt that he had adequate
evidence to justify his assumption that he could tell what the hybrids
were. Bees do work E. glauca vigorously, and if trees of
E. glauca and the citrus species were quite close together, I should
think there would be some cross-pollination.
At the same time, Dr. Furr notes in closing his discussion that: "Swingle
probably grew or had grown by his subordinates more seedlings of E. glauca
than anyone ever grew before or since his day."
BIGENERIC HYBRIDS
Eremolemons
[Eremocitrus glauca X Citrus limon 'Meyer lemon'].
All the bigeneric hybrids between
Eremocitrus and Citrus have unifoliolate leaves, intermediate in
size between the two parental species but thin and bifacial like those of
Citrus. Many years ago, all citrus trees except a Meyer lemon
were removed from the vicinity of a fruitful Australian desert lime in the
Coachella Valley at the U.S. Date and Citrus Station. Desert
conditions produced about one-fifth hybrids, probably with Meyer lemons as the
pollen parent. They grew vigorously, and when 8 to 10 feet high
flowered and fruited rather abundantly, producing subglobose, slightly oblate,
yellowish fruits, 18 to 25 mm in diameter, which have five or six segments and
usually one to three seeds. Upon germination, these seeds grew
rapidly and produced plants which looked exactly like the original
hybrid. Doubtless these eremolemon seeds contained only nucellar-bud
embryos which reproduced the mother F1 hybrid
exactly. This Coachella eremolemon, as it is called, has a high
tolerance for boron and doubtless a greater tolerance for salinity in the soil
moisture than any species of Citrus. Its seedlings are
remarkably uniform, grow rapidly when young, take Citrus buds readily,
and have taproots like the Citrus rootstock used by
nurserymen.
Eremoranges [Eremocitrus
glauca X Citrus sinensis].
Doubtless some of the many hybrids grown from seed from the Eremocitrus
trees at the U.S. Date and Citrus Station were of this
parentage.
Eremoradias [Eremocitrus
glauca X Citrus aurantium].
Perhaps
a few hybrids developed at the U.S. Date and Citrus Station were of this
parentage, as a sour orange tree grew only a dozen yards away from one of the
most fruitful trees of Eremocitrus glauca.
TRIGENERIC HYBRIIDS
Citrangeremos [Eremocitrus glauca X (Poncirus
trifoliata X Citrus sinensis)].
These hybrids showed trifoliolate or difoliolate leaves and usually also some
unifoliolate leaves. They were undoubtedly the result of the
pollination of Eremocitrus flowers from a row of citrange trees (Savage,
Morton, and Rustic) growing nearby in the U.S. Date and Citrus Station at
Indio. Many of these hybrids had very narrow leaflets, which
perhaps was the reason they were weak and of slow growth.
Both the bigeneric and the trigeneric hybrids of
Eremocitrus are very unlike any known citrus fruit trees, but evidently
they cannot properly be named as new genera and new species. If it
proves possible to cross-pollinate successfully a citrangeremo [=
Eremocitrus X (Poncirus X Citrus)] with another trigeneric
hybrid already existing, the faustrimedin [= Microcitrus X
(Fortunella X Citrus)], no fewer than five genera of True Citrus
Fruit Trees will have been combined in the resulting hybrid, viz.,
Citrus, Fortunella, Microcitrus, Eremocitrus, and
Poncirus! Naturally such ultra-complex hybrids, if obtained,
would not be recognized by taxonomists as constituting true genera or true
species.
XXIII. Poncirus
Raf.
XXIII. Poncirus Raf.
Sylva Tellur. Mant. 143. 1838. Pseudaegle Miq. Ann. Mus. Bot. Lugd.-Bat.
2:83. 1865-1866.
Type species
(monotype).—Citrus trifoliata L. = Poncirus trifoliata
(L.) Raf.
Distribution.—Central and
northern China; widely cultivated.
Common
name.—Trifoliate orange.
Small trees with
single, stout, axillary spines and palmately 3-foliolate, deciduous leaves,
twigs dimorphic; (a) normal twigs with internodes as long or longer than
the petioles, (b) foliage spurs, which develop from dormant buds on twigs
of previous year, with extremely short internodes (less than 0.5 mm long)
bearing 1-5 normal foliage leaves but no spines; flowers developing singly at
the nodes in spring from scale-covered flower buds formed early in the
previous summer on last year's twigs that lose their leaves during the winter;
sepals 5, petals 5, long, slender, spathulate or clawed; stamens 4 or more times
the number of petals, with free, glabrous, slender filaments; ovary subglobose,
pubescent, seated on a short, shallow, cup-shaped disk broader than the ovary;
ovary with 6-8 (sometimes fewer or more), usually 7 locules; ovules 4-8 in 2
rows in each locule; style short, thick, merging into the slightly clavate
stigma; fruits globose or short-pyriform, subsessile, 3-5 cm diam., finely
pubescent, with lemon-yellow, soft, orange-like peel with abundant oil glands;
pulp-vesicles elongate-conical, slender-stalked, bearing scattered, secretory,
hair-like organs, and filled with acid pulp containing numerous droplets of
acrid oil (or oily wax) in the center; seeds oval, plump, often with several
supernumerary nucellar embryos; germination hypogeous; young seedlings
developing at first bract-like cataphylls, then intermediate forms that soon
merge into normal 3-foliolate leaves.
This
remarkable genus, although evidently closely related to Citrus,
Fortunella, Microcitrus, and Eremocitrus, has many strikingly
aberrant characters. In the first place, it differs from all the
other True Citrus Fruit Trees, which are found in only tropical or subtropical
regions, in having penetrated far into the temperate zone in northeastern Asia;
in so doing it has become a deciduous tree with small leaf buds and larger
scale-covered flower buds (formed in early summer) that pass the winter on the
leafless terminal twigs and open before (and sometimes with) the leaves early
in the following spring. The trifoliolate leaves are doubtless a
survival of the foliage of some remote ancestral plant. All the
other True Citrus Fruit Trees have unifoliolate leaves (except Clymenia,
which has simple leaves) that persist on the tree for two or more years and
cannot endure severe cold.
The pith of the
stem shows transverse plates composed of thick-walled cells, not as yet found
in any other genus related to Citrus (see Swingle, 1909).
The pulp-vesicles carry scattered hair-like organs that bear at their expanded
tips a number of rounded, thick-walled cells with numerous small, oblique
fissures. These organs secrete a viscous fluid that allows the
pulp-vesicles to slip past one another. These organs are unknown in
any other genus of the orange subfamily. The immature fruits
contain ponciridin, a glucoside analogous to hesperidin, but not found in
Citrus.
Poncirus hybridizes
freely with Citrus. Such hybrids, called citranges,
citrandarins, etc., are usually ovule-sterile but occasionally produce some
fertile pollen grains. Poncirus has also been hybridized
(after overcoming many difficulties) with Fortunella and citranges have
been hybridized with Eremocitrus.
The
pulp-vesicles of Poncirus contain numerous droplets of oil.
Penzig (1887, pp. 161-62) discovered these inclusions and found that some of
these oil globules were semisolid and more or less brittle. Oil
droplets are also found in the pulp-vesicles of Citrus, being very
abundant in the species of Citrus which belong to the subgenus
Papeda.
Poncirus is not easy to
place in a phylogenetic series along with Citrus and other related genera
arranged so as to show the course of evolutionary progress.
Doubtless many "missing links" between it and the ancestors of Citrus
have perished, leaving Poncirus as perhaps the most isolated and
aberrant genus of all the True Citrus Fruit
Trees.
Poncirus trifoliata (L.) Raf. Sylva Tellur. Mant. 143. 1838.
Citrus trifoliata L. Sp. Pl. ed. 2. 1101. 1763; C. trifolia Thunb.
Fl. Japon. 294. 1784; C. triptera André (non Desf.), Rev. Hort.
57:518. 1885; Aegle ? sepiaria DC. 1824; Pseudaegle
sepiaria (DC.) Miq. Ann. Mus. Bot. Lugd.-Bat. 2:83. 1865-1866;
Limonia ? trichocarpa Hance, 1882. Illus. Kaempfer, Amoen.
Ext. 5:801. 1712; Hooker, Bot. Mag. pl. 6513. 1880; Nicholson, Gard.
Chron. 3 ser. 27:269. 1900; Penzig, Studi Bot. Sugli Agrumi, Atl., pls.
13, 14. 1887; André, Rev. Hort., 57:pl. facing p. 516 (col.). 1885;
Georgeson, Amer. Gard. 12:525. 1891; Swingle, U.S. Dept. Agr. Bur. Pl.
Ind. Circ. 46:figs. 1-6. 1909; idem, in Bailey, Stand. Cycl. Hort.
5:2751, 2752, figs. 3123-25. 1916; Tillson & Bamford, Amer. Jour.
Bot. 25:784, figs. 29-33. 1938; fig. 3-41 this work.
Type.—Based on Kaempfer's description and
plate.
Distribution.—Central and
northern China; widely cultivated in Japan, Europe, and North
America.
Common name.—Trifoliate
orange.
Small, much-branched tree, twigs of
rapid growth, strongly angled, becoming cylindrical with age; spines single,
stout, straight, sharp, oval in cross section (vertical axis longer); twigs
dimorphic: (a) normal twigs with internodes 1-5 or even 8 cm long with a
single leaf at each node, in the axil of which is a bud and often a stout
spine; (b) foliage spurs, arising from the nodes of the twigs of the
previous year's growth (but above the spine and 5-8 mm above the scar of
the fallen leaf in the axil of which it was laid down as a bud), with 1-5
(usually 2-4) excessively short nodes (less than 0.5 mm long), each bearing a
normal foliage leaf but no spine, tipped with a bud 1-2 mm wide, covered with
green, overlapping bud scales with more or less ciliate margins (such foliage
spurs may persist for two [or more?] years developing 2-4 or more leaves each
year); leaves palmately 3-foliolate; terminal leaflet 3-6 X 1.5-2.5 cm,
shallowly crenulate, serrate above the middle, cuneate at the base, midrib
slightly raised on both surfaces and gradually expanding below near the base
for 2 or 3 mm into a pulvinoid swelling entirely included in the leaf blade and
not free except at the extreme base (i.e., no petiolule), lateral leaflets
2.5-5 X 1-2 cm, sessile, lower half wider, petioles 1-2.5 cm, wing 2-3 mm at
broadest part, tapering to the base; leaflets on leaf spurs very similar but
with petioles slightly longer in proportion to the length of the middle
leaflet; spines stout, straight, acute, 1-6 cm long, 3-10 mm diam. at base,
2-4 mm diam. in the middle, oval in cross section (vertical diam. nearly twice
the transverse in the lower third of the spine); flowers produced in spring on
twigs of previous year's growth from subglobose flower buds, with bud scales
usually ciliate on the margins, flowers single, nearly sessile, 5- (rarely 4-
or 6-) merous; calyx composed of 5 small, nearly free, ovate, cucullate,
persistent sepals, 8-10 X 3-4 mm; corolla composed of 5 white petals, 18-30 X
8-15 mm, flat when fully open, subspathulate, often narrowed into a claw at the
base, soon deciduous; stamens 20 or more (up to 60); filaments free, unequal in
length, slender, tapering gradually toward the tips; ovary subglobose, 2 mm
diam., pubescent, with 6-8 (usually 7) locules; ovules numerous, in 2 rows in
each locule; style short, 1-1.5 mm long, merging into a rather large,
subclavate stigma; fruits almost sessile, globose, ovoid or slightly pyriform
(sometimes with a blunt nipple at the tip set in a depression surrounded by a
ring-shaped furrow), 3-5 cm diam., dull lemon-colored and fragrant when ripe,
finely and densely pubescent, peel 5-10 mm thick, with numerous oil glands,
rather rough; seeds ovoid, plump, very numerous, leaving very little space for
the pulp, which is composed of slender-stalked, narrow, elongate-fusiform
pulp-vesicles filled with very acid juice and having a slender column of
numerous small droplets of acrid oil in the center. The
pulp-vesicles bearing hair-like appendages, expanded at the tips; these usually
show scattered thick-walled cells having numerous small slanting fissures,
which secrete a viscous fluid.
The trifoliolate orange shows surprisingly few variations
considering that it has been grown in China for thousands of years and in Japan
since at least the eighth century. Palmately 4- or 5-foliolate
leaves are seen very rarely on old trees. In the parks of
Washington, D.C., a small-flowered form occurs that has shorter petals than
the normal form and these are occasionally seen bearing the rudiments of
anthers on their margins. In other words, such flowers show partial
staminody of the petals.
The curious dwarfed
variety with very narrow, almost threadlike leaflets, called hiryô in
Japan, where it is cultivated as a dwarf potted tree, is described below. The Comte de Castillon (1877, p. 73,
figs. 11, 12) reproduced Japanese figures and gave Latin varietal names to two
other sports of this species grown in Japan—the variety microcarpa,
which has minute fruits, and the variety punctata, which has leaves
dotted with golden-yellow spots, called in Japan sunago or "gold-dust"
trifoliate orange. These interesting garden sports probably do not
merit recognition in taxonomic botany as varieties.
In spite of the variations mentioned above, it is
probable that Poncirus trifoliata, of all the True Citrus Fruit Trees, is
the most stable of the species that have been cultivated for long periods by
man. This may perhaps be due to the fact that this aberrant genus
is not very closely related to the other genera in the group; if it crosses
with them, the resulting hybrids (citranges, etc.) are almost always
sterile. Thanks to this circumstance P. trifoliata has
preserved its nature with little change in spite of long culture.
The trifoliate orange has been used very
extensively in Japan and somewhat less so in the United States as a rootstock
for the satsuma and other cultivated citrus fruit trees. It is also
used occasionally for hedges (Swingle, 1911) and is commonly grown as an
ornamental in Asia, Europe, and North America, especially in regions that are
too cold to permit the culture of ordinary citrus fruit trees out of
doors.
Poncirus trifoliata has been
crossed with Citrus making hybrids called citranges that have proved
valuable as vigorous, hardy, disease-resistant rootstocks for ordinary citrus
fruit trees.
The Chinese have for centuries
used the dried fruits of the trifoliate orange in their medical
practice. Apparently both the immature fruit, called
chih-shih,and the fully ripe, sliced, and dried fruits, called
chih-k'o, are utilized, but for different
purposes.
Poncirus
trifoliata var. monstrosa
(T. Ito) Swing. in Bailey, Stand. Cycl. Hort. 5:2952. 1916. Citrus
trifoliata var. monstrosa T. Ito, Encycl. Jap. (Nippon Hyakka
Daijiten) 2:1056. 1909.
Type.—Cultivated in Japan.
Common
name.—Flying-dragon trifoliate orange.
Tokutaro Ito's original diagnosis in English reads as follows: "Citrus
trifoliata var. monstrosa T. Ito, nov. var. Branches and
spines crooked and curved, leaves very slender." A very dwarf
variety, with tortuous slender branches with curved spines; the leaves are very
small with the leaflets often reduced to linear filaments on which the oil
glands appear as node-like thickenings.
In
Japan this curious monstrosity, called hiryô, or flying dragon, is
esteemed for culture as a dwarf potted plant.
PONCIRUS
HYBRIDS
Citranges
[Poncirus trifoliata X Citrus sinensis].
As the genus Poncirus is monotypic, all
its hybrids are intergeneric, i.e., either bigeneric or trigeneric.
The first adequately safeguarded hybrids of Poncirus and Citrus
were made by Swingle in 1897. These hybrids, called citranges,
showed such striking differences, even between sister hybrids grown from a
single fruit, that a number of them (Rusk, Willits, Morton, Coleman, Savage,
Rustic, and Saunders) were named, described, and illustrated. (See
Swingle and Webber, 1898, p. 400, text fig. 13; Webber and Swingle, 1905, pp.
223-35, pls. 10-16, text figs. 12 and 13; Webber, 1907, pp. 329-36, pls. 17-20;
Swingle, 1910, pp. 36-41, pls. 1-2; idem, 1913e, pp. 83-87, pls.
on pp. 84, 86, 88, 90, 92; idem, 1913f, pp. 382-86, figs. 1-4;
idem, 1927b, pp. 19-21.) Several named varieties of
citranges are described by Hodgson in this work.
Citrumelos
[Poncirus trifoliata X Citrus paradisi].
Citrumelos are very similar to
citranges. At least one of them, the Sacaton, produces large
numbers of seeds containing only nucellar embryos and consequently yields
identical F1 seedlings in great numbers, like
the Troyer citrange (see Hodgson, this work). The Sacaton is a promising
rootstock.
Citrandarins [Poncirus trifoliata X Citrus
reticulata].
Citrandarins are much like
citranges except that the mandarin orange parent greatly reduces the amount of
bitter oil found in droplets inside the pulp-vesicles. In
citranges this oil usually makes the fruit too bitter to eat.
Citremons [Poncirus trifoliata
X Citrus limon].
Many of these
hybrids show abnormally small leaves in germinating and soon die (Swingle,
1913f); others are broad-leaved. One, the Highgrove, is a
very fast grower with fruits like a lemon but rougher and more
seedy.
Citradias [Poncirus trifoliata X Citrus
aurantium].
These hybrids greatly resemble
citranges but seem to be even more vigorous and cold-resistant. The
Brownell citradia (fig. 3-42) grows vigorously and fruits freely in northern Alabama
at Tuscaloosa (Lat. 33° 12’ N.). It makes a good stock for the
commonly cultivated citrus fruits.
Citrumquats [Poncirus trifoliata X Fortunella japonica, or
F. margarita].
These hybrids are very
difficult to make, germinate badly, and nearly all die while still very
small. Nevertheless, a very few of them have been obtained that grow
vigorously (see under Fortunella).
SECOND-GENERATION CITRANGES
Segentranges [(Poncirus trifoliata X Citrus sinensis)
F2].
Almost
all Poncirus X Citrus hybrids are completely ovule-sterile,
although almost all of them produce a little fertile pollen. Only
two of the many citranges that have been studied (the Phelps and the Sanford)
regularly produce true second-generation seedlings, all of which show
extraordinary variability. Some of the seedlings are almost like
the one or the other parent species, and all degrees of intermediates
occur. In the experimental plantings these hybrids have been called
segentranges, for convenience, but this name covers such a range of forms as to
be of little practical value (see Swingle, 1910, p. 37, pls. 3, 4 [showing
Sanford citrange F2 seedlings]; Swingle,
1927b, p. 20; and Hodgson, this work).
BACK-CROSSES OF PONCIRUS HYBRIDS ON THE PARENT
SPECIES
Citrangors [Citrange X Citrus sinensis].
These hybrids are often very like the sweet
orange in leaf characters; the fruits have not as yet been studied.
The orange-like seedlings resemble also the orange-like second-generation
seedlings of the fertile citrange varieties. Some of the citrangors
show trifoliolate leaves and look much like citranges (see Swingle, 1910, p.
36, pls. 7, 8).
Cicitranges [Citrange X
Poncirus trifoliata].
These hybrids
often look much like Poncirus trifoliata in both leaf and fruit
characters. The fruits are often more juicy and contain fewer seeds
than those of P. trifoliata but are always more or less bitter from the
droplets of acrid oil in the pulp-vesicles. All the cicitranges are
trifoliolate, but some are citrange-like with broad lateral
leaflets.
TRIGENERIC PONCIRUS HYBRIDS
Citrangequats [Citrange X Fortunella sp.].
These vigorous and variable hybrids are discussed
in this chapter under Fortunella.
Citrangedins [Citrange X (Fortunella X Citrus)
'Calamondin’].
A remarkable hybrid of this
parentage is discussed in this chapter under Fortunella.
Citrangeremos [Citrange X Eremocitrus glauca].
These hybrids are discussed in this chapter under Eremocitrus.
CITRANGES AND OTHER PONCIRUS HYBRIDS IN DOORYARD AND VILLAGE
GROVES
All Poncirus hybrids are
marked by their trifoliolate or difoliolate leaves and hence are easily
recognized even when the seedlings are too young to have flowered or
fruited. There is every reason to expect that similar spontaneous
hybrids due to cross-pollination by insects between Citrus and
Poncirus should be found; as a matter of fact, such hybrids do occur in
China, Indo-China, and Malaya. Also it is to be expected that
intergeneric hybrids between Poncirus and widely different species of
Citrus should exist; in fact, these are found in the homeland of citrus
fruits, in southeastern Asia, Japan, and throughout the Monsoon region.
Swingle saw in the dooryard gardens in China
trifoliolate citrange trees and also trifoliolate, difoliolate, and
unifoliolate forms that were probably citrangors; other forms, obviously
Poncirus hybrids, have been found in China and in
Indo-China. It is therefore to be expected that back-crosses and
second-generation seedlings, as well as complex 3-parental hybrids, should occur
not uncommonly as "chance seedlings" in dooryard citrus gardens in the Far
East.
Fortunately the vast amount of carefully
safeguarded hybridization work that has been done in the United States on the
True Citrus Fruit Trees has shown what types of hybrids may be expected to
occur, and as a matter of fact do occur, when mixed plantings of species of
Citrus, Fortunella, and Poncirus in dooryard or village
groves are cross-pollinated by insects (which visit the flowers with great
assiduity because of their abundant supply of nectar). Chance
seedlings are almost certain to arise from seeds dropped in such dooryard or
village groves and their chance seedlings can and do persist, flower and
fruit. If the fruit is valued, such hybrids are propagated by
grafting and a new cultivated variety is born.
Knowing, as we do, the exact origin of the remarkable hybrids of Poncirus
listed above, as well as of the many other, even more complicated, hybrids
listed under Fortunella and under Eremocitrus, we are able to appreciate how unscientific it would
be if "new species" (and even the "new genera" that would be needed in some
instances) were created to include these hybrids (whether they arose
artificially by human design or spontaneously through insect pollination), thus
giving them the same taxonomic status as genuine species that arise in nature
by slow evolutionary processes.
XXIV. Clymenia
Swingle
XXIV. Clymenia
Swingle, Jour. Arnold Arbor. 20:251.
1939.
Type species
(monotype).—Citrus polyandra Tan. = Clymenia polyandra (Tan.)
Swing.
Distribution.—Known only from
the type locality near Namatanai in northern New Ireland, Bismarck Archipelago,
northeast of New Guinea.
Common
name.—Clymenia.
This genus is allied to
Citrus; it differs in having (1) subsessile pulp-vesicles (somewhat
narrowed at the base but not borne on slender stalks) attached in great numbers
to the lateral segment walls for 3/4 the distance from the peel to the axis;
(2) very numerous stamens (50-100) with free, slender filaments; (3) ovary with
a very short, stout style (less than 1/2 the length of the ovary); and (4)
leaves with very short petioles, from 1/10 to 1/20 the length of the leaf
blade, with which they are not articulated. The flowers arise
singly in the leaf axils and are borne on straight, rather stout pedicels,
slightly longer than the petioles of the subtending leaves; calyx persistent,
5-lobed, lobes reflexed at apex in dried material, suberect at sinuses, thick
and coriaceous, wrinkled without, glabrous within; corolla 5-merous, petals
imbricate, dotted with numerous small oil glands; stamens very numerous (10-20
times as many as the petals), filaments free, slender, anthers oblong; ovary
ovoid with 14-16 locules, ovules several in each locule; style very short
(about half the length of the ovary); stigma capitate; disk broader than the
ovary, about half as long; fruit ovoid, size of a small orange, skin thin,
orange-like, dotted with oil glands; pulp-vesicles attached in part to the
dorsal, but principally to the side walls of the segments; seeds very numerous,
small, subglobose or oval, glabrous, embryo green. Small trees,
branches spineless, twigs subangular when young, then cylindrical; leaves thin,
smooth, with numerous subparallel lateral veins, acuminate-caudate at apex,
cuneate at base, tapering into the very short petiole, furrowed above, and not
articulated with the leaf blade.
This
remarkable new genus (of which only one species is known) is in its general
aspect so much like Citrus that it was at first mistaken for a sweet
lime. It bears sweetish edible fruits about the size and shape of a
sweet lime (5 to 6 by 7 to 8 cm), even to a short nipple at the
apex. The ripe fruits are yellow in color, again like sweet
limes. However, Clymenia differs very widely from
Citrus in many important taxonomic characters and is undoubtedly a
distinct new genus. It is especially noteworthy in having a type of
pulp-vesicle not known in any other citrus fruit.
The type of this genus, Citrus polyandra,
was described in Latin rather fully by Tanaka in all its characters except the
detailed structure of the pulp-vesicles and the manner of their attachment in
great numbers to the side walls of the fruit segments. The
pulp-vesicles are short, plump, blunt, oval or subglobose, sessile or very
short-stalked. What is also important from a taxonomic standpoint
is that these pulp-vesicles are attached in very large numbers all along the
side walls of the segments, nearly to the inner angle touching the central axis
of the fruit. The leaves of Clymenia are very unlike those
of any other True Citrus Fruit Tree (Citrus, Microcitrus,
Eremocitrus, Fortunella,or Poncirus) and strongly resemble
those of Monanthocitrus, a genus of the Minor Citroid Fruit Trees
described from New Guinea by Tanaka. Monanthocitrus has,
however, a much simpler floral organization with only twice as many stamens as
petals, instead of 10 to 20 times as many, as in Clymenia.
The fruits of Clymenia are about the size of a small orange.
The greatly enlarged disk bearing very numerous stamens is unique in the orange
subfamily.
Clymenia polyandra (Tan.)
Swing. Jour. Arnold Arbor [sic] 20:253. 1939. Citrus polyandra
Tan. Studia Citrol. 2:158, 163. 1928; Citrus medica subsp.
limonum var. limetta Lauterb. (non "[Risso] Engl.") Bot. Jahrb.
55:264. 1918. Illus. Tillson & Bamford, Amer. Jour. Bot.
25:786, figs. 34-36. 1938; Swingle, loc. cit. pl. 1, figs. 1-5; fig.
3-43 this work.
Type.—Bismarck
Archipelago, New Ireland, Namatanai, Nukonoku (Peekel, No. 408).
Herb. Bot. Mus., Berlin-Dahlem. Fragments of type: Herb.
Natl. Arbor., Washington, D.C., sheet No. 46011. Topotype:
New Ireland, Buratamtabai, Namatanai (C. S. Gee, 5/6/37, with the help of
Father Peekel!), Herb. Natl. Arbor, Washington, D.C., sheet No.
46010.
Distribution.—Bismarck
Archipelago: known only from the vicinity of Namatanai, New Ireland.17
Common
name.—Clymenia.
Tanaka's original
description (in Latin) may be translated about as follows: "Branches
triangular, glabrous, without spines; leaves oblong, caudate-acuminate,
abruptly narrowed at the base, irregularly serrate; petioles very short, not
articulated [with the leaf blade], and not winged, but furrowed [on the upper
side]; flowers single in the axils of the leaves, with rather long, somewhat
thick, pedicels; calyx saucer-shaped, obscurely toothed; stamens up to 100,
free, arising in many rows outside the disk; filaments linear; anthers oblong;
ovary ellipsoid; style thick, very short; stigma capitate, somewhat pentagonal;
fruit oblong, somewhat lemon-shaped, with an inconspicuous mammilla at the tip;
peel thin, segments numerous; pulp vesicular, yellow, rather sweet;
pulp-vesicles short, blunt, very numerous; seeds obovoid, superposed."
The twigs are spineless and the leaves are very
peculiar, being elliptical, acuminate at both ends, with the base merging into
a very short, slender, wingless petiole which is not articulated or in any way
marked off from the blade. The flowers are produced singly in the
axils of the leaves, petals short and broad (7 X 7 mm); stamens very numerous
(50-100) in many whorls, with short filaments (5 mm), pistils shorter than the
stamens, ovary ovoid, with a broadly rounded tip not merging into the style,
ovary with 14-16 locules, style short (only half as long as the ovary), with
capitate stigma; fruit lemon-shaped, 4.5 X 6-7 cm, with a short nipple at the
apex; pulp sweet; seeds numerous, superposed, polyembryonic.
Tanaka deserves the credit for having first
noticed the great divergence of this plant in many of its essential taxonomic
characters from any other of the known True Citrus Fruit Trees.
However, he considered it to be a hybrid of Citrus macroptera and C.
medica, which is said by Lauterbach to be "commonly planted at the [European
?] stations."
After studying Citrus
macroptera and other members of the subgenus Papeda and observing the
hybrids between them and species of the subgenus Citrus that are common
in the Philippines, Swingle became convinced that a hybrid such as Tanaka
thought his Citrus polyandra to be (Citrus macroptera X C.
medica) could not possibly show a wingless petiole, even though one supposed
parent, the citron, has an almost completely wingless petiole, because the
other supposed parent, C. macroptera, has a very long, very broadly
winged petiole (certainly the largest petiole to be found in Citrus).
The extraordinarily close resemblance of the
leaves and petioles of this species to those of Monanthocitrus convinced
Swingle that this plant was an entirely new type of citrus fruit tree, possibly having descended from a remote ancestral species common
also to Monanthocitrus. Swingle thereupon published
Clymenia as a new genus to include this plant (see fig. 3-43).
This species,
called a-mulis by the natives in the village of Namatanai in New Ireland,
is cultivated for its sweet fruits.
XXV. Microcitrus
Swingle
XXV. Microcitrus
Swingle, Jour. Wash. Acad. Sci. 5:570.
1915.
Type.—Citrus australasica
F. Muell. = Microcitrus australasica (F. Muell.)
Swing.
Distribution.—Of the 6 known
species, 5 are found in eastern Australia (Queensland and northern New South
Wales) and the other (M. warburgiana) in southeastern New
Guinea.
Common name.—Australian wild
lime.
Twigs minutely puberulous; leaves
dimorphic, those on young seedlings greatly reduced cataphylls gradually
merging into small, linear or elongate-elliptical leaves and finally into the
normal foliage of the species; mature leaves more or less coriaceous, strongly
veined, glabrous except in midrib near base; petioles short, wingless, finely
pubescent, especially on the upper flattened side; spines slender, acute,
single (or in pairs in M. inodora and M. maideniana) in the axils
of the leaves; flowers small, 4-5-merous, stamens entirely free, ovary with 4-8
locules; ovules several, 4-8 or more in each locule; fruits round-ovoid or
cylindric; pulp-vesicles noncoherent, slender-stalked, subglobose or ovoid
(except in M. australis, in which they are elongate-fusiform, tapering
above), filled with acid pulp having minute droplets of acrid oil aggregated
along the axis; seeds small, ovate in outline; cotyledons hypogeous in
germination.
This genus is obviously related
to Citrus, from which it differs, however, in having dimorphic foliage,
free stamens, an ovary with four to six (or eight) locules, and more or less
coriaceous, strongly veined leaves. Moreover, it is related to the
highly specialized genus Eremocitrus, which also has dimorphic foliage
and free stamens. Eremocitrus, however, differs from
Microcitrus in always having thick, coriaceous leaves with thick cuticle
palisade tissue and stomata on both faces and an ovary with only three to five
locules having only two ovules in each locule.
The six known species of Microcitrus are
the result of millions of years of slow evolution from a primitive ancestral
type. From this ancestral type arose the genus Eremocitrus,
with marked xerophytic adaptations and two ovules in each locule of the ovary,
and the genus Microcitrus, with more numerous ovules in each locule.
This ancestral type probably resembled somewhat
M. warburgiana, the New Guinea species, which has small leaves and small,
nearly spherical fruits. From such an ancestral form, one line of
evolution produced the so-called native orange, round lime, or Dooja (M.
australis), that grows to a large tree and has subglobose fruits much larger
than those of M. warburgiana, with long, slender, pointed, more or less
twisted pulp-vesicles; another line of evolution culminated in M. inodora
and M. maideniana, highly specialized forms showing adaptations to
tropical rain forests, with large leaves and paired spines; a third line of
evolution led to the small-leaved, somewhat xerophytic species M.
australasica and M. garrowayi, both with long-ovoid or very
elongate-cylindric fruits.
These remarkable citrus fruits are extremely interesting, in that
they show how evolution has proceeded in regions isolated as Australia and New
Guinea have been during the last twenty or thirty million years since they were
cut off from all other land masses.18 The evolution of
other citrus fruits is not so easily followed, since Citrus,
Fortunella, and Poncirus did not originate in regions that were
geographically isolated in definitely dated geologic eras. A key to
the species of Microcitrus is presented.
1. Microcitrus australasica
(F. Muell.) Swing. Jour. Wash. Acad. Sci. 5:572. 1915. Citrus
australasica F. Muell. Fragm. Phytogr. Austr. 1:26. 1858. Illus.
G. W. Oliver, U.S. Dept. Agr. Bur. Pl. Ind. Bul. 202:pl. 15, figs. 5,
6; pl. 2, fig. 1. 1911; F. M. Bailey, Comprehen. Cat. Queensl. Pl. 84, fig. 63.
1913; Swingle, loc. cit., figs. 1-4; fig. 3-44, E this
work.
Type.—Moreton Bay, Queensland (F.
von Mueller). Present location
(?).
Distribution.—Australia: coastal
regions, southern Queensland; northern New South
Wales.
Common name.—Australian
finger-lime.
First leaves of young seedlings
small, 3-5 X 2-4 mm, ovate, with very short wingless petioles borne on numerous
slender, horizontal twigs with internodes often only 2-4 mm long, with sharp,
slender spines, 7-12 mm long, at the nodes; the upright twigs of young plants
have similar leaves and spines but the internodes are sometimes 5-8 mm long;
the upright, strongly angled, vigorous twigs of mature trees, which have
internodes 8-12 mm long with single sharp spines, 5-10 mm long, have
lozenge-shaped leaves, 22-25 X 14-15 mm, obscurely toothed in the upper half,
gradually narrowed to a blunt, truncate base with which the petiole
articulates; lateral veins faintly marked above, more clearly below, 6-10
pairs, arising at an angle of 35°-45° with the midrib, lower ones longer, upper
ones soon branching, all with several anastomosing curved cross-veins; leaves
tapering into a truncate or emarginate tip, those on lateral twigs 8-20 X 2-10
mm, the smaller ones abruptly truncate, often irregularly emarginate; spines
slender, short, 3-8 mm long; flowers arising singly in the axils of the leaves
(or occasionally in pairs); flower buds subglobose or obovate, 4 X 5 mm, borne
on very short pedicels, 1-2 mm long; flowers usually 5-merous but sometimes
3-4-merous; sepals free, small, concave, minutely ciliate; petals oblong, 7-8 mm
long; stamens 20-25, free, pistils short, stout, ovary with 5-7 locules; ovules
numerous, 8-16 (or even 20) in each locule; fruits cylindric-fusiform, 6.5-10 X
1.5-2.5 cm, often slightly curved, narrowed at both tip and base, often showing
a blunt protuberance at one or both ends; peel rough with numerous oil glands,
greenish-yellow at maturity; pulp-vesicles nearly free or loosely cohering,
long-stalked, ovoid, ending in a very blunt or rounded tip, about 1.7-3 X
1.2-1.5 mm, borne on a slender stalk 1-2 mm long; seeds numerous, small, 6-7 mm
long, ovoid, monoembryonic, usually flattened on one side and often showing
small, shallow depressions on the other faces (probably caused by mutual
pressure of the pulp-vesicles exerted during development of the fruit); testa
smooth (not wrinkled as in Eremocitrus glauca); cotyledons hypogeous in
germination.
This, the type species of the
genus Microcitrus, has several characters separating it from all of the
species of Citrus. It is also very clearly separated from the
other species of Microcitrus by its very long, slender fruits that are
unique in the orange subfamily (see fig. 3-44,E). They have numerous seeds (or
rudiments) in each segment (more numerous than in any other true citrus
fruit). The pulp is composed of loosely grouped, long-stalked,
subglobose or pyriform pulp-vesicles, tapering bluntly to the tip.
These pulp-vesicles are very different from those of M. australis but
are much like those of M. inodora; they also resemble somewhat those of
Citrus (Papeda) hystrix, as noted by Penzig (1887, pp.
214-17), who was the first to describe the anatomy of the fruits, seeds, and
pulp-vesicles of M. australasica. The very acrid pulp has a
harsh aftertaste, probably due to droplets of acrid oil in the pulp-vesicles,
such as have been found in the Australian round lime. Francis
(1929, p. 174) stated that the fruits, both of this species and of M.
australis, "have a lemon-like flavor accompanied by a taste of a turpentine
nature."
The finger-lime of the coastal
regions of northeastern Australia is a tall shrub or small tree and makes a
handsome ornamental. The seedlings are very spiny. The
first leaves are minute linear cataphylls; these gradually merge into juvenile
foliage which, in turn, merges into the mature foliage, the leaves of which are
smaller than those of any other True Citrus Fruit Tree with the exception of
Eremocitrus glauca when the latter occurs in very dry
situations. The branches on young plants often stand out nearly at
right angles to the stem, giving the young plant the appearance of a very
dwarfed fir tree. This species is precocious; Oliver (1911, p. 15)
figured a small branch of a two-year-old seedling bearing two fruits.
Uphof (1932, pp. 138-39), in an examination of
five trees of M. australasica, found that most of the flowers were male
and that only a very few were perfect. He examined the flowers on
eight twigs and found, out of 161 flowers, that 151 were male and that only ten
were perfect.
Microcitrus australasica
is of interest in hybridization work because of the great vigor and the
extraordinary production of slender leafy twigs that have been shown by the Sydney hybrid.
1a. Microcitrus australasica var. sanguinea (F.
M. Bail.) Swing. Jour. Wash. Acad. Sci. 5:574. 1915. Citrus
australasica var. sanguinea F. M. Bail. Dept. Agr. Queensl. Bot. Bul.
18:8. 1892. Illus. Penzig, Studi Bot. Sugli Agrumi, Atl. pl. 21,
fig. 13. 1887.
Type.—Queensland (Bailey
?). Herb. Bot. Gard. Brisbane.
Distribution.—Australia: southeastern
Queensland.
Common name.—Red-pulp
finger-lime.
This variety differs from the
species in that the pulp-vesicles at maturity vary from pink to red in
color.
There are similar "pink" or red
varieties of oranges (the so-called "blood oranges") and grapefruit, some of
which are known to have arisen as budsports. However, the
red-pulped variety of the Australian finger-lime is found growing wild and can
be propagated from seed; it seems to have originated without the aid of
man.
HYBRIDS
Faustrimedins
[Microcitrus australasica X (Fortunella sp. X Citrus sp.)
'Calamondin'].
This trigeneric hybrid is
discussed under Fortunella.
2. Microcitrus australis
(Planch.) Swing. Jour. Wash. Acad. Sci. 5:575. 1915. Limonia
australis. A. Cunn. nomen subnudum. 1839; Citrus australis
Planch. Hort. Donatensis 18. 1854-1858; C. planchoni F. Muell. 1872.
Illus. Penzig, Studi Bot. Sugli Agrumi, Atl. pl. 21, fig. 8-13. 1887;
Campbell, Agr. Gaz. N. S. Wales 10:1168. 1899; Guilfoyle, Austr. Pl. pl.
facing p. 123. 1911; Bailey, Comprehen. Cat. Queensl. Pl. 84, fig. 62. 1913; fig.
3-44,A-B this work.
Type.—Moreton Bay, Queensland (Leichhardt). Herb. Mus. Hist.
Nat., Paris.
Distribution.—Northeastern
Australia.
Common name.—Australian
round lime.
Leading terminal twigs straight,
very angular, with internodes 1-1.5 cm long, with single slender acute thorns
5-10 mm long; lateral twigs short, slightly angled, with internodes 5-10 mm
long, and spines 2-6 mm long; leaves entire, glabrous, or sparingly ciliate on
the margin near the base; leaves articulated with the petioles; larger leaves
on leading twigs 3-4 X 2-3 cm, obovate to obcordate (sometimes lozenge-shaped
on less vigorous leading twigs); leaves of lateral twigs diamond-shaped, 2-3 X
1.2-1.8 cm; veins visible on both surfaces, but more conspicuous below, 12-15
pair arising at an angle of 25°-30° with the midrib and frequently branching,
with veinlets arising at a very acute angle and running parallel to the veins;
leaves rounded, emarginate or bluntly pointed at tip, cuneate at the base;
petioles articulated with the leaf blades, finely pubescent and flattened an
the upper side, 2-3 mm long; flowers arising singly in the axils of the leaves,
small, 10-15 mm diam., 4- or 5-merous, with 16-20 stamens with free filaments;
fruit globose or subglobose, 2.5-3.5 or 5 cm diam., with 6 segments (Schaler's
green to grass green; Ridgway, 1912, pl. 6); pulp-vesicles proper (exclusive of
stalk) 6-10 X 2.5-3.5 mm, lightly coherent in mature fruits, very pale greenish
(water-green, Ridgway, pl. 41) near the peel but much brighter green (light
grass-green to grass-green, Ridgway, pl. 3) in the center, irregularly
slender-pyramidal or fusiform, more or less angular from pressure, tapering
gradually into long, more or less contorted blunt tips, 1-1.5 mm wide,
containing numerous large droplets of strong-flavored oil, stalks often very
short, sometimes 2-3 mm long, texture of pulp-vesicles firm enough to cut into
slices with a sharp knife (much firmer than in any species of Citrus);
seeds flattened, containing a single embryo; no clearly defined chalazal cap in
spite of the acid nature of the pulp (in Citrus the species with acid
pulp have strongly colored chalazal caps).
This species, called "Dooja" by the aborigines and round lime or native orange
by the Australians, is native to northeastern Australia. It is
readily distinguished from all the other species of Microcitrus (and from
Eremocitrus as well) by its globose or slightly pear-shaped,
rough-skinned fruits, 2.5 to 5 cm in diameter (about the size of a large
walnut), and by its short-stalked pulp-vesicles that taper gradually into blunt
tips often more or less deformed and twisted by mutual pressure (see fig. 3-44,B).
The early
descriptions of this species and of M. australasica are so meager as to
leave doubt concerning which species was being discussed.
Fortunately the original descriptions of Citrus australis, published by
Planchon in a rare work (Hortus donatensis, 1854-58, p. 18), was based on
a specimen in the herbarium of the Muséum d'Histoire Naturelle at Paris,
collected by Leichhardt in the vicinity of Moreton Bay in southeastern
Queensland. A study of this type specimen makes clear the proper
application of the name M. australis.
Penzig (1887, pp. 210-17, pl. 21, figs. 8-13) described in detail the anatomy
of this species. He found that the pulp-vesicles were similar to
those of the commonly cultivated citrus fruits except that they contain, along
their central axis, large masses of yellowish oil. These
pulp-vesicles are very different in shape from those of the finger-lime, M.
australasica, or those of the other species of Microcitrus.
The Australian round lime, or Dooja, grows to be
a tree 30 to 60 feet high and is the largest of any of the native citrus fruit
trees. The fruit is said to be used for preserves in
Queensland. This species, which has been introduced into the United
States, grows well in California, where it is now fruiting. It is
exceeded in vigor, however, by the Sydney hybrid, the result of a natural cross
of this species with the finger-lime, Microcitrus
australasica.
HYBRIDS
Sydney
hybrid19 [Microcitrus australis
X M. australasica]. Illus. fig. 3-44,C and D.
Twigs very slender and very numerous, usually
only lightly angled, very minutely and rather sparsely puberulous; bud scales
brownish, inconspicuously ciliate; nodes 5-13 mm long, with a single slender,
sharp, axillary spine, 4-12 mm long; leaves on vigorous leading twigs 20-30 X
8-13 mm, glabrous, narrowly elliptical or lozenge-shaped, with undulate or
irregular, shallowly crenate-dentate margins, narrowed at the apex, sometimes
with an emarginate tip, cuneate at the base, with numerous lateral veins
arising at a small angle (25°-35°) with the midrib, with some cross and some
subparallel veinlets, narrowed into very short petioles, 2-3 mm long which are
sparingly and finely pubescent above; leaves on lateral twigs 10-20 X 3-8 mm,
linear-elliptic or narrowly lozenge-shaped, the smaller leaves entire, the
larger with undulate or irregular, shallowly dentate margins, petioles 1.2-2 mm
long; fruits elongate-obovoid or ellipsoid, 35-50 X 20-28 mm, with the tip
abruptly rounded, numerous slightly protuberant oil glands, circular in outline
and about 0.5 mm diam., peel 2 mm thick, yellowish-green when ripe (near lime
green, Ridgway, pl. 31); inner portion 0.4-0.5 mm thick, smooth, of firmer
texture than the outer, segments 5-8 central core solid, very narrow, ovule
traces numerous; pulp-vesicles ovoid, tapering to an acute point above, broadly
rounded at base, 4.5-7 X 2-4 mm, borne on slender stalks, 2-7 mm long, attached
to inner wall of the ovary but not to segment walls, not adhering to one
another, containing an acid juice; seeds none. The ripe fruits have
a spicy, not disagreeable odor. The flower buds are spinel red, and
the new growth is very dark dull dusky purple (Ridgway, 1912).
This remarkable hybrid was grown from seeds of
the Australian round lime, Microcitrus australis, or of the finger-lime,
M. australasica, sent to the U.S. Department of Agriculture's
Agricultural Research Service by the late J. E. Maiden, director of the
Botanical Garden at Sydney, Australia. More than 200 meters of
twigs, both large and small, were borne on a single branch, 3 cm in diameter, of
a Sydney hybrid growing near Riverside, California, which survived for several
decades on land no longer irrigated where citrus trees made little or no
growth.
The leaves are intermediate in shape
between the two parent species, but, owing to the enormous number of twigs and
consequent great profusion of leaves, they are not intermediate in size between
those of the parent species, but nearer that of the small-leaved parent, the
finger-lime. The leaf margins are more crenate than those of either
parent. The small twigs of both parents are minutely ciliate, but
the larger angled twigs, especially of the finger-lime, are almost
glabrous. In the hybrid, however, the twigs are more pubescent than
those of either parent and are more slender and more upright.
This hybrid has been found to support the orange
and other commonly grown citrus fruits very well in greenhouse culture, and it
can be propagated readily from soft-wood cuttings of summer growth.
As it excels all known citrus fruit trees in the length of twigs it produces,
it should be tested as a stock for citrus fruits grown on foothill soils low in
organic nitrogen. On account of its extremely abundant branchlets
covered with miniature leaves, the Sydney hybrid deserves trial as an
ornamental, especially in the orange-growing section of the southeastern
states.
3. Microcitrus garrowayi
(F. M. Bail.) Swing. Jour. Wash. Acad. Sci. 5:574. 1915. Citrus
garrowayi F. M. Bail. Queensl. Agr. Jour. 15:491. 1904. Illus.
Bailey, Comprehen. Cat. Queensl. Pl. 84, fig. 65 (leaves and fruit).
1913.
Type.—Queensland, Mount White,
Cape York Peninsula, near Coen (Lat. 13° 55' S., Long. 145° E.), altitude
1,300 feet (R. C. Garroway). Herb. Bot. Gard.
Brisbane.
Distribution.—Known
onlyfrom the type locality.
Common
name.—Garroway's Australian wild lime.
Bark with minute, irregularly interrupted longitudinal ridges; young twigs very
slender, 0.8-1.5 mm diam., cylindric or rarely slightly angled when very young,
with very small more or less upward curving, hyaline hairs; internodes 4-15 mm
long (usually about 6-10 mm), with very slender spines, 2-5 mm long, or
spineless; older twigs below last growth show gray, corky, longitudinal, more
or less confluent streaks, 0.5-1 mm wide, that show fissures (these corky
streaks become more and more abundant and finally run together, so that
two-year-old twigs show little or no green bark); leaves of cotype specimen
(from Mount White, Queensland) narrow, thick and leathery, lozenge-shaped to
broadly lanceolate, 30-45 X 15-20 mm (small leaves 20-25 X 10-12 mm), bluntly
pointed, rounded or even slightly emarginate at apex, cuneate at base, with 5-7
pairs of veins, visible on both surfaces of the leaves, arising at the midrib
and occasionally branching, margins nearly entire, slightly undulate; leaves on
leading twigs of greenhouse specimen 20-25 X 12-18 mm, lozenge-shaped or
broadly ovate, bluntly rounded or slightly emarginate at apex, cuneate at base,
margins slightly dentate or crenulate especially toward the apex, with a few
hyaline hairs on the margin at the base of the leaf and still fewer on the
lower portion of the midrib; petioles 2-4 X 0.8-1 mm, wingless, flattened above,
articulated with the blade, finely pubescent on the upper flattened side,
especially at the edges; fruit of cotype specimen ellipsoid or
cylindric-ellipsoid, 4 X 2 cm, with numerous sunken oil glands.
Bailey described this species as follows, under
the name of Citrus garrowayi: "Fruit upon the slender branchlets oblong,
2 1/2 in. long, 14 lines diameter. Oil glands large, giving a
tuberculose appearance to the fruits; ultimately these glands sink, and the
fruit appears then to be lacunose; rind very thin; cells 4 or 5; pulp of a
sharp agreeable acid; seeds 3-angular, white, free, with more or less very
short hairs, about 3 lines long and 2 lines thick in the centre.
The rough rind of the fruit somewhat resembles that of C. australis, but
the form of fruit is nearer to that of C. australasica; the fruit of this
latter, however, is nearly smooth and the glands
small."
Microcitrus garrowayi seems to
be a good species, distinct from M. australasica and M. australis,
occurring farther to the south in eastern Australia. Bailey called
this the Mount White lime. It has been introduced into the United
States.
4. Microcitrus
inodora (F. M. Bail.) Swing. Jour. Wash. Acad. Sci. 5:577. 1915.
Citrus inodorus F. M. Bail. in Meston, Rept. Govt . Sci. Exped.
Bellenden-Ker Range 34. 1889; C. inodora F. M. Bail. Syn. Queensl. Fl. 3d
Suppl. 12. 1890. Illus. Bailey, Queensl. Fl. 1:pl. 10. 1899;
idem, Comprehen. Cat. Queensl. Pl. fig. 64. 1913; C. T. White, Jour. Hered.
13:120, figs. A-E. 1922.
Type.—Queensland, Harvey's Creek, Russell River (F. M. Bailey).
Herb. Bot. Gard. Brisbane; part of type material in Herb. Natl. Arbor.,
Washington, D.C.
Distribution.—Australia: northeastern
Queensland.
Common name.—Large-leaf
Australian wild lime.
Leaves broadly oval or
lanceolate, or with cuneate bases and more or less acute or even caudate at the
apex, 8-18 X 4-10 cm, with very numerous, parallel, lateral veins arising at
an angle of 60°-80° with the midrib; petioles very short, 4-8 mm long,
wingless, not articulated with the blade; flowers small (as in other species
of Microcitrus) but reported by Bailey to be odorless; stamens free but
much more numerous (over 30!) than in the other species of Microcitrus;
fruit oblong or elliptical (somewhat lemon-shaped), 5-6.5 X 3-3.5 cm, with a
conical base, segments 8, on drying sometimes showing ribs corresponding to the
walls separating the segments; pulp-vesicles stalked like those of other
species of Microcitrus, rather loose; seeds small, somewhat pear-shaped,
6-8 X 4-5 mm.
This species is a shrub or small
tree 2 to 4 meters high with the trunk 4 to 5 cm in diameter. The
twigs are angular like those of Citrus sinensis (the sweet orange) and
have one or two slender, very sharp spines, 6 to 12 mm long. It has
been found growing wild only along Harvey's Creek, Russell River, where,
according to White (l.c., p. 119), it is common in the lowland rain forests at
the foot of the Bellenden-Ker Range in northern Queensland near Cairns (Lat.
17° S.). This is a very rainy region, the average annual
precipitation being 170 inches. In February, 1922 (?), the rainfall
was 62 inches!
5. Microcitrus
maideniana (Domin) Swing. Jour. Wash. Acad. Sci. 28:533. 1938.
Citrus maideniana Domin, Bibl. Bot. 89:297. 1927. [Beiträge z. Fl.
Austr. 851].
Type.—Northeastern
Queensland, rain forest along Harvey's Creek (Domin, December 1909, to January
1910). Herb. Univ. Praha,
Czechoslovakia.
Distribution.—Known
only from the type locality.
Common
name.—Maiden's Australian wild lime.
The
original diagnosis reads, in approximate translation: "A low erect shrub,
1.5-2.5 m high; young twigs angular-compressed; spines numerous on the twigs,
two in the axil of each leaf, of which one spine is occasionally shorter;
leaves lanceolate or ovate-lanceolate, pale green, midrib rather prominent;
fruit borne on a short thick pedicel, obovoid-ellipsoid, when ripe about 4.5 cm
long and 2.5 cm wide or sometimes less, yellow, smooth, with 8 obtuse ribs, not
clearly visible in ripe fruits, apex deeply sunken; young fruits deep green
with closely set dusky green spots.
"A new
species named in honor of the late Professor J. H. Maiden of Sydney; it appears
from the description to be related to Citrus inodora F. M. Bail. from the
same locality, but, besides the smaller fruits, the deeply sunken apex shows it
to be distinct."
It is highly probable that
this species is very closely related to Microcitrus inodora, native to
the same locality, with which it agrees in having paired spines and more or
less clearly eight-ribbed fruits (doubtless having eight segments).
The deeply depressed apex of the fruit is the only clearly distinctive
character known as yet.
6.
Microcitrus warburgiana (F. M. Bail.) Tan. Bul. Soc. Bot. France
75:714. 1928. Citrus warburgiana F. M. Bail. Contr. Fl. Brit. N.
Guin. 1. 1903. Illus. Bailey, loc. cit. first unnumbered plate,
lower half (leafy twigs and fruit); fig. 3-45 this work.
Type.—Southeastern British New Guinea, Milne Bay (W. E. Armit).
Paratype: U.S. Natl. Herb., Washington, D.C. (from F. M.
Bailey!).
Distribution.—Known only from
the type locality.
Common name.—New
Guinea wild lime.
The original description
reads as follows: "Branchlets angular, soon becoming terete, apparently quite
glabrous; axillary spines erecto-patent, about 2 lines long on the specimen
seen. Leaves rhomboid-lanceolate, 1 1/2 to 2 1/2 in. long, 5 to 11
lines broad, deeply emarginate and the margins crenulate for 2/3 the way down;
primary nerves rather numerous and very oblique, the intermediate ones and
reticulate veins rather prominent. Petioles light colored, 2 to 3
in. long, very narrowly winged. (No flowers collected.)
Fruit axillary on a stout peduncle, 1 1/2 lines long, globose, about 3/4 in.
diam., oil glands concave, crowded, rind thin, cells 6. This
species seems to be nearly allied to C. Medica, var. aruensis,
Warbg., Engl., in Bot. Jahrb., 1891, page 340."
A twig of the type collection sent to Swingle by
Bailey shows characters as follows: Twigs 30 cm long, 2 mm diam. at base, with
very slender terminal twigs only 0.5-1 mm diam., internodes 1-2.5 cm long
(rarely 5 mm on the slenderest lateral twigs), slightly angled at first but
soon cylindrical, minutely puberulous at first but apparently soon glabrescent;
leaves elongate-elliptical, leaf blades 4-6 X 1-1.8 cm, coarsely but shallowly
crenulate-serrate, bluntly rounded at apex and often slightly emarginate, bases
cuneate, subentire, lateral veins 10-12, connected by numerous smaller
subparallel reticulating veinlets; petioles subulate, very short, 2-3 mm long,
0.5 mm thick, 0.8-1 mm wide, articulated with the leaf blade, with slightly
depressed channel on the upper side bordered along the upper half by a slightly
crenulate membrane (in drying, sometimes curved upward and making a nearly
closed channel), sparingly and minutely puberulous on the upper side and on the
margins.
The small globular fruits, the
elongate-elliptical, crenulate leaves, and the narrowly subcrenulate-margined,
very short petioles separate this New Guinea species very clearly from the
other species of Microcitrus. This is the only species of the
genus outside of Australia, and it occurs nearly a thousand miles away from
M. garrowayi, the Australian species of Microcitrus growing
nearest to New Guinea.
The fruits are small,
globose, with six segments (fig. 3-45). Bailey's description, however, says
nothing concerning the character of the pulp-vesicles, if any! The
petioles differ from those of the other species of Microcitrus in having
a very narrowly crenulate margin or wing. The veins are numerous,
making only a small angle (35° to 50°) with the midrib, and soon branch,
forming a reticulation of veinlets not unlike those of M. garrowayi,
although the latter species has smaller leaves with fewer veins.
The ultimate twigs, especially the short lateral branchlets, are remarkably
slender, the smallest lateral twigs being 1 to 1.5 mm or even only 0.5 to 0.8
mm in diameter.
XXVI. Citrus
L.
XXVI. Citrus L. Sp. Pl.
1:401. 1753. Citrophorum Neck. Elem. 1:401. 1790;
Sarcodactylis Gaert. f. Fruct. Sem. 3:39, pl. 185. 1805;
Papeda Hassk. Flora, 25, Beibl. 2:42. 1842.
Type species.—Citrus medica
L.
Distribution.—Southeastern Asia,
East Indian Archipelago, Philippines, New Guinea, New Caledonia, Bismarck
Archipelago, Solomon Islands, Fuji (?), Samoa (?).
Small trees; young twigs angled, soon
cylindrical, with single spines in the axils of the leaves but older branches
often spineless; leaves 1-foliolate, usually thin, not coriaceous, lateral
veins few and without conspicuous reticular veinlets between; petioles usually
more or less winged and articulated with the leaf blade (except in C.
medica, where they are wingless or merely margined and not articulated with
the leaf blade); flowers single in the axils of the leaves or in short,
axillary, corymbose racemes, perfect or staminate by more or less complete
abortion of the pistil; calyx cup-shaped, with 4-5 lobes; petals 4-8 (usually
5), thick, linear, strongly gland-dotted, imbricate in the bud; stamens usually
about 4 times as many as the petals but in some species usually 6-10 times as
many; disk annular, short; ovary subglobose and sharply distinct from the much
narrower style, or else truncated, fusiform, or subcylindrical and merging
gradually into a style nearly as thick as the upper portion of the ovary,
locules 8-18 (usually 10-14), with 4-8 or more ovules in each locule in 2
collateral rows; style cylindrical, expanding abruptly into the subglobose or
oblate-spheroid stigma; fruit a hesperidium with the segments containing seeds
near the inner angle and the rest of the space filled with stalked fusiform
pulp-vesicles, filled with a very watery, large-celled tissue; around the
segments is a white endocarp, outside of which is the peel dotted with very
numerous oil glands and turning yellow or orange at full maturity; seeds
obovoid or flattened obovoid, more or less angular, containing 1 or many
embryos, either white or green.
In young developing fruits of Citrus short-stalked,
club-shaped slime-secreting organs develop rapidly along with the
pulp-vesicles. These, however, soon mature and liberate, from
collapsing superficial cells of the head, slimy matter which probably enables
the developing pulp-vesicles to slide freely over one another (see Tschirch and
Oesterle, 1898, p. 303, pl. 70, figs. 36, 36a, 36b, 36c,
and 38). Similar organs probably occur in the other genera of the
True Citrus Fruit Trees, but in the subgenus Protocitrus of the genus
Fortunella these slime-secreting organs are sessile and are not intermixed with
the developing pulp-vesicles.
The genus
Citrus, thanks to the progress made in the taxonomic study of the orange
subfamily in the last half century, can now be seen in proper
perspective. It is the culmination of a very long period of
progressive evolution that certainly began before Australia was cut off from
land connection with New Guinea and Asia, probably more than 20 million years
ago. The genus Microcitrus, closely related to
Citrus, occurs both in southern New Guinea (one species) and in
northeastern Australia. A related genus, Eremocitrus,
originated in Australia and became a xerophyte adapted to grow in semiarid
regions. Citrus occurs in New Caledonia, but only one species
is known there or in neighboring islands to the east and southeast.
The genus may have originated in the New Guinea-Melanesia region, but its
evolution into many different species took place chiefly on the mainland of
southeastern Asia. In fact, it is only there that the most highly
developed species of Citrus can be considered as indigenous.
Only the aberrant species belonging to the subgenus Papeda have developed
in the East Indian Archipelago, and in the Philippines, New Guinea, and
Melanesia. A few of these species reached the Asiatic mainland, and
from these sour and bitter-flavored, almost inedible species have doubtless
developed the fragrant, delicious-flavored species of the subgenus
Citrus, in many ways the finest fruits known to man.
The superiority of these finer species of
Citrus has led to their extinction as wild plants because their fruits
made them so attractive that the trees were transplanted to village
gardens. This led to cross-pollination by bees and other insects,
with the resultant obscuring of species limits to the extent that in many parts
of southeastern Asia and in the East Indian Archipelago it is difficult to find
any clear-cut species of Citrus. There appears instead, an
endless array of complex hybrids, often combining not two, but three or even
four, different species in varying proportions. This situation has
led to much confusion in attempting to define species of Citrus.
As will be shown later, it is necessary to abstain from naming the principal
cultivated varieties of Citrus as independent species.
Unfortunately, names have sometimes been given even to almost sterile varieties,
doubtless of hybrid origin, that depend wholly on man's care for their
perpetuation.
THE SPECIES CONCEPT IN THE GENUS CITRUS
About a century and a half ago, several
treatises were published on the citrus fruit trees. The first of
these was the Traité du Citrus, by Georges Gallesio (1811), followed by
the work of J. A. Risso (1813) and J. L. A. Loiseleur-Deslongchamps and Etienne
Michel (1818), and finally by the Histoire naturelle des orangers of
Risso and A. Poiteau (1818-1822). In these voluminous works, every
cultivated variety was given a name consisting of a Latin phrase of from three
to ten, or even more, words. As these polynomials did not fit into
the Linnaean system of plant nomenclature then in universal use by botanists,
they were never incorporated into the taxonomic literature, especially since
botanists considered that hybrids and monstrosities originating in orange
houses or gardens were not deserving of technical botanical Latin names.
The view of professional botanists on this
plethora of names is perhaps best indicated by quoting a paragraph relating to
the genus Citrus written by Daniel Oliver. In his monograph
on all the other genera of the orange subfamily occurring in India, Oliver
(1861, p. 1) gave his reasons for not attempting to treat the species of
Citrus. He wrote: "Feeling it utterly hopeless usefully to
define them [the species of Citrus], I have thought it best to leave the
Oranges, Lemons, Limes and their allies as I found them. It has
been difficult to form, for purposes of comparison, any satisfactory
approximate estimate of the number of species in this very variable and widely
cultivated genus. From the data afforded by Risso, Loureiro, Wight
and Arnott, Miquel, and others, and from what I have myself seen of dried or
living specimens, I have assumed five as about the probable number."
Fourteen years later, in 1875, J. D. Hooker in
his Flora of British India included Citrus among the thirteen
genera that he recognized as comprising the orange subfamily, but recognized
only four species in Citrus, in India proper, Ceylon, and Burma!
As late as 1896, Adolph Engler, who for many
years had specialized on the Rutaceae, recognized only six species of Citrus
for the entire world, and in these he included three species now placed in
other genera. Finally in 1931, as his last and greatest taxonomic
contribution, Engler revised his account of the Rutaceae for the second edition
of Die naturlichen Pflanzenfamilien. In this he included
eleven species in the genus Citrus and some six or seven species (which
he had placed in Citrus in 1896 but now considered to be distinct) in the
two genera Poncirus and Microcitrus.
In the meantime, early in the present century,
evidence of the most convincing sort, based on many hundreds of accurately
safeguarded cross-pollination experiments made by the citrus experts of the
former Bureau of Plant Industry of the U. S. Department of Agriculture, began
to be published. This evidence showed that all the species of
Citrus tested, and also species of two other related genera, could be
hybridized without difficulty and that these hybrids often manifested
astonishing variability in the first generation. What was even more
surprising, many of these hybrids, especially those between rather remotely
related species, bore apparently normal seeds that reproduced the parent hybrid
exactly. In reality it was found that such hybrids often produced
no viable egg cells, but, if pollinated, tiny buds from the nucellar tissue of
the mother plant would grow into the embryo sac and there develop into
apomictic embryos that, of course, reproduced the mother plant exactly (Traub
and Robinson, 1937).
It is therefore highly
probable that in China, Japan, India, and other Oriental countries accidental
hybridization has occurred in dooryard or village plantings of assorted citrus
fruit trees through the action of bees and other insects which seek the
abundant nectar of Citrus flowers. It is also reasonable to
suppose that if hybrids did arise and if chance seedlings came into fruiting
they would be propagated by the villagers if the trees bore fruit of superior
quality.
Many year of intimate contact with
the citrus hybrid populations gave Swingle and his colleagues a clear
understanding of the characters and nature of some of the more valuable of
these hybrids between different species of the commonly cultivated citrus fruit
trees. Armed with this experience, Swingle made three trips between
1915 and 1926 to the citrus-growing regions of Japan, China, and the Philippine
Islands, where he paid special attention to hybrids and mutations of pummelos,
oranges, and other citrus fruits. He recognized in Oriental
villages many of the types of hybrids that had been made in the United States
by carefully safeguarded cross-pollinations! In the Orient, these
had originated as accidental hybrids, doubtless due to cross-pollination by
insects. For example, citranges, hybrids of the trifoliate orange
and the sweet orange, were found growing in dooryards in the Yangtze Valley
region of China, and tangelo-like fruits, undoubtedly hybrids of pummelos and
loose-skinned citrus fruits, were not uncommon in Japan! After the
mass of evidence regarding the origin and nature of citrus hybrids was brought
to light and published in detail in well-illustrated papers, evidence that
proved the possibility of producing large numbers of striking new forms of
citrus fruit trees by cross-pollination of the commonly cultivated species of
Citrus, it is indeed surprising that A. W. Lushington in India, in 1910,
and T. Tanaka in Japan, from 1924 to date, should have given species rank to
scores of citrus forms that undoubtedly are hybrids.
Tanaka (1935b, p. 576) realized that many
of his "secondary or miscellaneous" species are chance hybrids. In
discussing the fifteen so-called "primary elements or species" of
Citrus, Poncirus, and Fortunella,20 and the "secondary or
miscellaneous species derived from them," he stated that the lack of secondary
species among the pummelos (Citrus grandis) of southern China, "is to be
explained by the lack of cooperation of compatible elements (species) acting as
the mating pair to originate new species."21 Actually, one of
the conditions that hinders the hybridization of C. grandis with other
species is that the anthers lie against the stigmatic surface and release their
pollen before the bud opens. Later Tanaka stated: "Compatible
combinations of primary species22 encourage the creation of
such secondary elements…of great significance to the evolution of the Citrus
flora of the world."
In 1932 Tanaka
(1932d, p. 263) made the following statement concerning the Calamondin
orange: "The admittance [acceptance] of Citrus microcarpa [= C. mitis, a distinct bigeneric hybrid between Citrus and
Fortunella] is another of the endless examples, which present no
contradiction to the writer's system of nomenclature…" Tanaka also
stated explicitly (1927a, p. 54) that new forms of Citrus to
which he had given taxonomic rank as genuine new species might have arisen
from "chance seedlings." He considered that the Japanese
Natsudaidai, which he held to be a good species (C. natsudaidai Hayata),
was "unquestionably a chance seedling originated at Hagi in Yamaguchi
Prefecture." He also considered the satsuma orange to be
"botanically a good species, Citrus unshiu Marc.," that had originated
some centuries earlier in western Japan as a chance seedling of one of the
famous oranges of Hwangyen, Chekiang Province, China. In discussing
the cultivated Ichang lemon (probably a hybrid of the Ichang papeda, C.
ichangensis Swing., with some other species of Citrus grown in
central China), which he named C. wilsonii (1932b, p.
37). Tanaka stated: "The present species unquestionably has
Citrus junos in one parent and seems to be a cultigen originated through
the chance seedling." There is no proof, however, that any one of
the chance seedlings discovered and propagated by man as cultigens could have
persisted and become established as a wild species.
The taxonomic delimitation of species,
subspecies, and forms in Citrus and closely related genera is complicated
by three peculiarities in the reproduction of citrus fruit trees: (1)
polyembryony through formation of nucellar-bud embryos, (2) rejuvenation by
neophyosis of such nucellar-bud embryos of more or less senescent varieties
long propagated asexually, and (3) the spontaneous production of autotetraploid
forms.
Hybrids of Citrus, both
intergeneric and intrageneric, are frequently egg-sterile; yet, when
pollinated, they often set seeds. These seeds are devoid of
sexually derived zygotic embryos ("gametic" embryos of some authors), but
contain one or more embryos within the embryo sac apomictically produced by
ingrowing buds arising from the adjoining undifferentiated nucellar tissue of
the female parent. This phenomenon was first described by
Strasburger (1878) and later was discussed by Frost (1626) and Traub
(1936). Such nucellar-bud seedlings reproduce the mother plant
exactly, so that many violent F1 Citrus
hybrids, such as citranges (C. sinensis X Poncirus trifoliata)
and tangelos (C. paradisi X C. reticulata), reproduce perfectly
from seeds and sometimes are grown on a large scale. Taxonomists
have found such egg-sterile hybrids to be confusing, since they do not show the
usual wide variation in the second generation characteristic of normal
F2 seedlings but behave like very constant true
species.
In many species of Citrus,
such nucellar embryos may occur together with the zygotic embryo.
In such cases, the zygotic embryo may produce a plant that varies somewhat in
its characters from the mother plant, whereas the nucellar embryos reproduce
the mother variety almost exactly. Nucellar-bud seedlings as well
as sexual seedlings apparently restore the variety to its primitive or juvenile
condition, because such plants not only develop spines abundantly but show
greater energy of growth and greater hardiness and sometimes produce somewhat
larger or otherwise superior fruit. Two nucellar strains of a
well-known mandarin orange (Dancy tangerine) were named the Weshart and Trimble
tangerines by Webber and Swingle (1905, pp. 238-40 and pl. 20-22).
The Silverhill satsuma is another striking example of a nucellar seedling of
the Owari satsuma (see Swingle, 1931).
Similar
rejuvenation of nucellar seedlings was observed in experimental work with the
Washington navel orange where there could be no confusion between the zygotic
and the nucellar embryos arising from the same seed, since the zygotic embryos
were citranges obtained through pollinations with Poncirus
trifoliata. This rejuvenation of the old cultivated varieties of
Citrus has been called neophyosis (see Swingle, 1932, 1933).
Nucellar strains of old varieties rejuvenated through neophyosis might easily
be mistaken for taxonomically distinct varieties or even subspecies, because
of their vigorous growth, their large leaves and fruit, and their luxuriant
development of spines which are often lacking on varieties long propagated
vegetatively.
Finally, thick-leaved seedlings
of several species of Citrus were found by Frost (1925, 1926) to be autotetraploid forms of the parent species;
the fruits have a thicker rind with larger oil glands, are usually more
depressed-globose, and have less juice, and more and larger seeds than the
diploid parental species from which they originated.
As a matter of fact, from the standpoint of
phylogenetic taxonomy, which records the evolutionary descent of natural wild
species, only Tanaka's "primary elements or species" deserve consideration as
true species; chance seedlings, mutations, and hybrids, propagated usually by
grafting, should be given a status similar to cultigens of apple or rose
varieties that no taxonomist of standing has proposed to name as good species,
although they often exhibit important differences in flowers or fruits or other
organs as well.
Once this nomenclatorial
confusion is cleared up, Tanaka's studies of the myriad cultivated races of
Citrus and Fortunella in the Far East will be appreciated at its
full value and he will receive recognition as having greatly advanced the study
and classification of citrus fruits. This task can then be carried
on without upsetting the scientific taxonomy of Citrus and
Fortunella, which is based on the native wild species originally
propagated spontaneously from seed without human help.
A. E. Kozhin, the well-known Russian
citrologist, published a notable paper on the citrus fruit (1931) in which he
gave particular attention to the very extensive literature on the subject and
to the taxonomic history of Citrus and the genera most closely related to
it. He strongly objected to the practice of considering "garden
species" as species in the taxonomic sense. He contrasted such
false species, some of them seedless and unable to live at all without man's
help and most of them propagated by clones, with true species. He
said in part: "…a Linnean species is a whole system of elementary forms which,
continually crossing with one another, keep within the limits of definite
specific characters. In drawing up such a system of forms, the
geographic moment, i.e., the conditions of habitat, play an important role…From
this point of view the attempt to introduce into the system as equivalent
members (as has been done by Tanaka), the wide-compassed Linnean species and
such narrow forms as garden species, frequently seedless ones, is a
failure."
More recently Stebbins (1950)
discussed the species concept in agamic complexes, and stated that "…the
species concept…maintained by the majority of those who desire a truly
biological concept of species, centers about the possibility for exchange of
genes between members of the same species and separation of different species
by barriers to the exchange of genes.…Such a concept cannot be applied to
agamic complexes. Free exchange of genes between apomicts is
prevented by the very nature of their type of reproduction, while the origin of
many apomictic clones is from genotypes which have combined the genes of
previously isolated sexual species, and which without apomixis would not be
able to persist because of their sexual sterility. It is not
strange, therefore, that systematists have not been able to agree on the
boundaries of species in…[apomictic genera]." Stebbins continued:
"In attempting to set up species like those in sexual groups, they are looking
for entities which in the biological sense are not there. Nor are
those 'splitters' who make a separate species out of every apomictic
clone…likely to provide any better concepts of the variation patterns in the
genera concerned.…One criticism is that the number of apomicts in any
well-developed complex is so large that recognizing them as separate species
makes the comprehension of the group as a whole difficult or impossible; one
cannot see the forest for the trees! But more important is the fact
that in many complexes the great majority of the apomicts, and in nearly all
complexes at least some of them, are only partly or facultatively
apomictic. From time to time they reproduce sexually, and on such
occasions a whole series of new clones, or 'species', may arise in the offspring
of a single individual." Once "splitting" is begun on the basis of
fruit characters or small detail, it would be possible to justify thousands of
species in the genus Citrus. Furthermore, this number would
be constantly augmented by additional "species" as the citrus breeder produces
new apomictic hybrids each year. As Lawrence (1951) has pointed
out, the presence of apomixis makes possible the survival of hybrids that would
otherwise be eliminated under natural conditions by their
sterility. Lawrence stated, "The perpetuation of these apomictic
hybrids has resulted in some descriptive taxonomists treating each biotype as a
morphologically distinct and seed producing species."
Two major systems of citrus taxonomy and
nomenclature have developed. Swingle's system is presented
here. The other is the system proposed by Tyôzaburô Tanaka
(1927c, 1935a, 1954, and 1961), and supported by Yuichirô Tanaka
(1948). Each taxonomic treatment is based upon years of research
and study by very competent men of widely divergent viewpoints.
Swingle began the first known attempts at
artificial hybridization of citrus in 1893. His experience as a
citrus breeder and his observations of large seedling populations produced by
controlled cross-pollination lead to the development of his system of
classification. He assembled an extensive collection of
Citrus material for study upon which he based this system.
Reece, the junior author, who was given the task
of revising this chapter for this edition of The Citrus Industry, makes
no pretense of competence in the field of taxonomy. On the other
hand, he has devoted many years to Citrus breeding. He
considers that Swingle's system comes closest to the truth of Citrus
taxonomy, and he cannot accept the Tanaka system with its excessive
splitting.
In following up his interest in
Citrus taxonomy, Tanaka has traveled widely and observed a wide range of
citrus. He is familiar with both the original literature and
herbarium material. Swingle and Tanaka became associated and
collaborated together at various times between 1915 and 1930 in this country and
Japan. Nevertheless, their divergent viewpoints are apparent in
their systems and conclusions. Their disagreement is confined
principally to the species problem in the genus Citrus.
Although Tanaka in 1935 recognized only fifteen
primary species in Citrus, Poncirus and Fortunella, he also
stated, in an English summary of a Japanese paper (Tanaka, 1935a): "The
author [Tanaka] enumerated 74 species as such well-established species of
Citrus, Fortunella and Poncirus in the present paper, but
the mobilization of all species from various Citrus districts is now on the way
of progress." In a later paper (1937), he listed no fewer than four
additional "species" of Lushington (published in 1910) and described six new
species himself, as well as four new varieties, all from India. In
several papers since 1935, he mentioned en passant many other new
species or varieties without describing them. This brought the
total to about 100, but the end was by no means in sight.
Tanaka (1954) established the foundation for his
classification system with the publication of Species Problem in
Citrus. In that work, he classified the forms of Citrus
into two subgenera, eight sections, thirteen subsections, eight groups, two
subgroups, two microgroups, and 145 species. Seven years later
(Tanaka, 1961), his system was expanded to include two new subsections,
Eulimonellus and Megacarpa, one new group, Paranobilis, and
twelve new species. Further refinements have continued, however, and
a classification chart sent by Tanaka to some of his colleagues in 1966
contains five new subsections and includes 159 species. In contrast
to the Tanaka system, the Swingle classification recognizes two subgenera: the
subgenus Papeda containing six species, and the subgenus Citrus
(formerly Eucitrus) with ten species. A comparison of the
major classification systems of the genus Citrus is presented in table 3-3. Systematics are given for the Tanaka
classification of 1961, which contained 157 species, and the Swingle
classification of 1943 followed in this chapter. In addition, the
table includes columns for species recognized by the pre-Englerian, Englerian,
and early Swingle and Tanaka systems.
Tanaka
recognizes thirty-five species of mandarins. In some cases the only
distinguishing character in his key is leaf size or fruit size.
Anyone familiar with Citrus production knows that size is easily
influenced by rootstock, soil, amount of moisture, fertilization, and weather
conditions prevailing during development of a particular flush of growth.
Tanaka's arguments for granting species rank to
some of the mandarins are interesting and often quite unique. In
regard to making the satsuma a separate species from Citrus reticulata,
he says: "Pulp qualities are…the real object of human interest to consume the
fruit. The public knows more about such specific distinction than a
few technicians because they consume more. To the Japanese, who eat
500,000 tons of satsuma fruit a year, they would never believe it to be a mere
variety or clone of either the Ponkan (C. reticulata) or Kunenbo (C.
nobilis), as is true to Americans not admitting the grapefruit as a variety
of the pummelo (Shaddock). It is a human belief and science should
endorse it." (See Tanaka, 1954, pp. 20-21.)
In support of his argument that the Ponkan, the
Dancy tangerine, and the C. crenatifolia of Lushington are separate
species and not varieties of C. reticulata, he comments that "if these
three should belong to a single species, 350,000,000 Indian people would not
believe in science." However, the objective of science is to seek
the truth rather than the support of human beliefs. Nevertheless,
few biologists doubt that new species can result from hybridization, but the new
species should be able to perpetuate itself by sexual methods. It
should not be dependent upon man for some type of vegetative
propagation. For this reason a number of Tanaka's "species" are
not considered valid by the authors. Furr and Reece (1946) found
"Citrus temple and C. clementina of Tanaka" unable to reproduce
their type even by nucellar embryony because nearly all embryos are
zygotic. Reece and Childs (1962) in a study of a seedling
population of Persian lime (C. latifolia Tanaka) have shown that this
variety is apparently a hybrid between a seedy lime (C. aurantifolia
Swing.) and citron as the other parent. If, indeed, a seedy lime
and a citron were the parents of the Persian lime, the appearance of the many
lemon types of seedlings in the population supports Swingle's hypothesis that
the lemon may be of "hybrid origin, perhaps having the citron and the lime for
parent species."
Santiago (1962), in a guide
to Malayan nomenclature of citrus species, followed Swingle's system of
classification with a few exceptions. Bhattacharya and Dutta
(1956), in their comprehensive monograph on the citrus fruits of Assam, followed
Swingle's system in a general way, but accept C. nobilis Lour., C.
karna Raf., C. limetta (Risso) Lush., C. jambhiri Lush., and
C. megaloxycarpa Lush., five species of the Tanaka system that were not
accepted by Swingle. They find the nomenclature of one of these
Tanaka species invalid (C. pennivesiculata Tanaka = C. megaloxycarpa
Lush.), and they name one new species found in Assam (C. assamensis
Dutta and Bhattacharya, sp. nov.). This is known as
ada-jamir or the ginger citrus of India because people consider its aroma
similar to that of ginger. It is taxonomically close to C.
hystrix DC., but differs from it to an extent considered sufficient by these
authors to deserve a separate rank.
Several
other systems of citrus classification have been put forward by various
authors. Perhaps the one that has received the least attention by
contemporary students of Citrus taxonomy is the system proposed by
Marcovitch (1926), which bases classification on leaf and flower characteristics
only.
Wolfe (1959) has criticized both
Swingle's conservatism and Tanaka's system with its excessive
splitting. He takes an intermediate position and recognizes eleven
species not considered valid by Swingle and Reece. These species
are indicated in table 3-3 (see footnote). The authors' reasons for
their rejection of these species are stated elsewhere in the text under the
discussion of the several forms whose taxonomic rank is in dispute.
The late R. W. Hodgson made two trips to the
Orient and studied the principal Citrus collections in India for nearly a
year. He also studied a large number of Citrus introductions
from many parts of the world. He worked in close association with
Tanaka for nearly a year when Tanaka was in the United States as a Fullbright
research scholar. Hodgson (1961; Hodgson, Singh, and Singh,
1963a, 1963b, and 1963c) published his nomenclature of
Citrus which recognized twenty-three Citrus species of Tanaka's
system. Swingle's treatment was criticized by Hodgson on the basis
that it denies species standing to ancient, well-known, distinctive forms of
economic importance. However, such criteria are not sound reasons
for species standing if they are apomictic and show evidence of their hybrid
nature. It is true that Swingle's treatment may involve speculation
concerning the probable parentage and even error in proper placing of the form
in the attempt to arrive at a natural system of classification.
However, if they are made in an attempt to understand the natural relationships
of biological organisms, do such errors merit greater censorship than a more
artificial system that names as species forms that are known or probable
hybrids, obvious apomicts, or horticultural forms that can only be perpetuated
by vegetative propagation?
Distinctive or
easily recognized form is not an adequate reason for naming
species. The situation in Citrus is somewhat analogous to the
situation in Canis domesticus so well expressed by Dobzhansky (1955):
"The longer a species has been domesticated the more numerous and more
diversified are its domestic varieties. Thus the dog is the most
ancient domestic animal, and has produced the largest number of
breeds. Furthermore, some dog breeds differ so much in appearance,
as well as in temperament and behavior, that it is hard to believe that they all
belong to the same species. Compare the little Chihuahua with a
Great Dane; or an ebullient Fox Terrier with a stolid Saint
Bernard. Nevertheless, all these breeds do belong to one species,
since they are capable of exchanging genes, and often do so, either directly by
occasional hybridization, or via the intermediate breeds where crossing is
impossible because of extreme difference in size. The dog species
has a common gene pool, subdivided, to be sure, into the gene pool of the
various breeds." Zoological taxonomists consider that those
distinctive forms are "cultivated varieties" not different species.
Certainly the related forms or "breeds" of citrus (the species of Tanaka) have
a common gene pool since they can and do exchange genes through
hybridization.
The study of D. Singh, and C.
A. Schroeder (1962) on the taxonomic and physiological relationships of the
so-called mandarin-lime group of Citrus caused them to conclude: "…the
so-called mandarin-lime of Citrus is a misnomer in that lime parentage is
not indicated. The possibility of mandarin-rough lemon parentage is
suggested." They contend that the binomial Citrus limonia
Osbeck should be employed as a species designation for members of this
group. Determination of the actual parentage of this group is not
easy to establish, but hybrid origin is not only possible, but highly
probable. Apomixis strengthens this probability. The
percent of polyembryony in germinated seeds does not, however, indicate the
degree of apomixis in these varieties. It only indicates the number
of seeds in which two or more embryos were of sufficient size and strength to
emerge on germination and single embryos are also of apomictic origin in many
instances. On the basis of probable hybridity and apomixis, the
binomial of Citrus limonia as a species designation for this group is
denied.
The choice as to which of these
taxonomic systems to follow will depend upon the personal viewpoint of the
student of citrus. The biological facts in the case will not be
altered by arguments for or against either system. However, serious
difficulties arise when taxonomists rank apomictically perpetuated biotypes
with sexual species as Heslop-Harrison (1956) has pointed out. Some
taxonomists following such methods have described as species over 1,100 forms
in the hawthorn genus, Crataegus, over 4,000 in the blackberry genus,
Rubus, and over 10,000 in the hawkweed genus,
Hieracium. It would be a simple matter to embark upon a
similar path and describe an astounding number of Citrus "species," but
the multitude of binomials, unnecessarily complicated keys, and cumbersome
system of nomenclature would make the comprehension of the group difficult
indeed.
In the two decades since World War II,
truly remarkable progress has been made in describing the chemical
constituents in citrus tissue. Because of their great economic
importance and adaptability to industrialization, the common citrus fruits have
been the subject of more intensive biochemical investigation than perhaps any
other horticultural crop. As a result, hundreds of compounds,
particularly those found in fruits and seeds, have been isolated, identified,
and in many cases assayed (Braverman, 1949; Bartholomew and Sinclair, 1941;
Sinclair, 1961; U.S. Dept. of Agriculture, 1962). More recently
such research has been extended to citrus varieties and species of little or no
economic importance and also to some of the near relatives of citrus.
The material summarized in table 3-4 provides an example of this type of research and
indicates the great variety of flavonoids found in the genus Citrus and
some relatives. Similar research is in progress on essential oils
(U.S. Dept. of Agriculture, 1962; Stanley, 1963a; Scora, 1966; Scora,
et al., 1966; Scora and Newman, 1967; Scora and Torrisi, 1966), coumarins
(Stanley, 1963b), carotenoids (Yokoyama and White, 1965), limonoids
(Dreyer, 1965), and other compounds found in leaves, fruits, or seeds of the
Rutaceae family.
In time the pattern
distribution of some of these compounds may prove to be distinctive within
certain genera, species, or other groupings. Such chemotaxonomic
considerations, when added to the classic morphologic and cytogenetic bases of
classification, should contribute toward a more penetrating insight into the
species problem of citrus.
A key to the subgenera and species of Citrus is
presented.
Subgenus Citrus
Subgenus
Eucitrus Swing. in Webber & Batchelor, Citrus Indus. 1:395.
1943.
Pulp-vesicles with few and minute, or
sometimes no, oil droplets (never containing acrid oil); petioles wingless or
narrowly winged, or, if broadly winged, then never 3/4 as broad as the leaf
blades; flowers large and fragrant; stamens cohering in bundles.
The genus Citrus is divided into two very
distinct subgenera, Citrus and Papeda, easily distinguished by
leaf, flower, and fruit characters. The subgenus Citrus
includes all the commonly cultivated species of Citrus, all of which have
pulp-vesicles filled with pleasantly acid, subacid, or sweet juice, free, or
almost free, from droplets of oil. On the contrary, none of the
species of Citrus belonging to the subgenus Papeda have edible
fruits, as the pulp-vesicles have dense aggregations of droplets of acrid oil
that form an axile column and usually give the juice a very disagreeable,
acrid, bitterish flavor.
In the course of a comparative study of the vascular traces of
the flowers of all obtainable genera of the orange subfamily, Tillson (1938,
pp. 21 and 30; also Tillson and Bamford, 1938, pp. 788 and 790) brought to light
a striking difference between the species of Citrus belonging to the
subgenus Citrus and those classed here in the subgenus
Papeda. The commonly cultivated species of Citrus, the
sweet orange, the mandarin, the grapefruit, the lime, and the lemon, all
belonging to the subgenus Citrus, show "not only fusion of the sepal
midrib with the lateral petal bundles [as is usual in the tribe Citreae], but
also fusion of the lateral sepal bundles with the petal midrib.
When the petal midrib reaches its position beneath the petal base, one or more
branches from it continue into the base of the calyx, forming the lateral sepal
bundles." On the contrary, Tillson found that the species of
Citrus belonging to the subgenus Papeda showed no fusion of the
lateral sepal bundles with the petal midrib (this was true even of C.
ichangensis, which differs from the other species of the subgenus Papeda
in having large flowers, much like those of the species of the subgenus
Citrus). In this character of its floral anatomy the subgenus
Citrus differs not only from the subgenus Papeda, but also from
all the other genera, Fortunella, Eremocitrus, Poncirus,
Clymenia, and Microcitrus, all closely related to Citrus, that
comprise the True Citrus Fruit Trees.
1. Citrus medica L. Sp. Pl. 2:782. 1753. Citrus
tuberose Mill. Gard. Dict. ed. 8. 1768; C. odorata Roussel, Fl.
Calvados 194. 1796; C. fragrans Salisb. 1796; C. cedra Link, 1831;
C. cedratus Raf. 1838; C. crassa Hassk. 1844. Illus. Risso
& Poiteau, Hist. Nat. Orang. pls. 96, 97, 99, 100 (all col.). 1818-1822;
Bentley & Trimen, Med. Pl. 1:pl. 53 (col.). 1880; Swingle, in Bailey,
Stand. Cycl. Hort. 2:779, fig. 971. 1914; and many
others.
Type.—Wanting, but plant seen
and studied by Linnaeus.
Distribution.—China and India southward; cultivated in many subtropical
regions.
Common name.—Citron.
A shrub or small tree of irregular habit of
growth; twigs angled and purplish when young, soon cylindrical, glabrous, with
stout, short, single spines in the axils of the leaves; leaves glabrous,
elliptic-ovate or ovate-lanceolate, bluntly pointed or rounded at the tips,
cuneate or rounded at the base, margins serrate; petioles short, wingless or
narrowly margined, not clearly articulated with the leaf blade; inflorescences
short, few-flowered racemes; flower buds large, purplish; flowers perfect or
male by more or less complete abortion of the pistil; petals 5, pinkish on the
outside; stamens very numerous, 30-40 or even 60 as found by Webber (1923, pp.
112-20); ovary large, bulged, cylindrical, with 10-13 locules, tapering into
the thick style, which is sometimes persistent; fruit large, oblong or oval,
surface smooth or more often rough and bumpy, fragrant, yellow when ripe, rind
very thick, segments small, filled with pale greenish pulp-vesicles with acid
or sweetish pulp; seeds numerous, small, 9-10 X 4-5 X 3-4 mm, pointed at the
base, smooth; embryo white. (See Hodgson's description of the
cultivated varieties of citron in this work.)
The citron was the
first citrus fruit to reach the Mediterranean region. Apparently it
was introduced to the eastern Mediterranean area following the invasion of
Persia by Alexander the Great about 325 B.C. Theophrastus (writing
about 310 B.C.) called the citron the Median or Persian apple. He
said that it was inedible but very fragrant and a remedy for rheumatism and
sore mouth, as well as a repellent to drive away moths. Engler
stated (1931, p. 338): "Since the fruits had the same uses as the wood of the
Sandarak tree, Callitris quadrivalvis, the [ancient] name of this wood
'Citrus' was transferred to the fruit as Mala citrea." This
renaming of the Median apple as the Citrus apple led to the transfer of the
name "Citrus" first to the citron and later to other citrus fruits.
The native home of the citron has not been
determined with certainty. The citron is commonly supposed to be
indigenous to India but J. D. Hooker, who said (1875, p. 514) that he had no
doubt about the citron's being truly wild when he found it growing "mainly on
dry sunny slopes [in Sikkim] totally unsuited for any kind of cultivation…,"
later came to doubt its being indigenous. Bonavia (1888, p. 70)
stated: "I am still in doubt whether it [the citron] is indigenous in
India. It does not appear to have any ancient Sanskrit name and the
number of varieties, if they are variations, on the western seacoast is
suggestive. It is curious that they should be found in the area
which came most in contact with foreigners."
The citron has been grown since ancient times in China, but Chi Han, minister
of state under the Emperor Hui Ti, in a work written about 300 A.D. (Nan fang
ts'ao mu chuang) mentioned the arrival, in 284 A.D., as tribute to the
Chinese Emperor, of 40 Chinese bushels of citrons from Ta-ch'in (a name usually
meaning the Roman Empire). He stated: "…the Barbarians value the
citron very highly. It is aromatic and its flesh is very thick and
white…" This early Chinese record of the citron would indicate that
it was not indigenous to China but had been introduced from the West.
The early advent of the citron in Media and
Persia, and its subsequent slow penetration into India and China, could be
explained easily if the citron should prove to be a native of southern
Arabia. The bael fruit of India, Aegle marmelos, has no close
relatives in Asia, but three closely allied genera, Aeglopsis,
Afraegle, and Balsamocitrus, are found in Africa.
Citropsis, an African genus of the Near-Citrus Fruit Trees closely
related to the Asiatic genus Atalantia, has eleven species.
It would not be surprising to find midway between India and Africa, in some
mountain oasis within the tropical zone in Arabia, the citron growing in a wild
state. Over a century ago Wellsted (1838, vol. 1, pp. 126-52) found
gardens in the Jebel Akhbar Mountains (150 km southwest of Mascat) where grapes
grew abundantly; also "pomegranates, citrons, almonds, nutmegs and walnuts
with coffee bushes." Over thirty years ago Bartram Thomas (1932,
map, p. 101) explored thoroughly the Qara mountain range, about 1,100 km
farther to the southwest, and found it to be "an Arcadia of luxuriant forests
that clothe steep mountains with perennial streams." In these
mountains, situated in a summer rain belt along the coconut-fringed shores of
the Arabian Sea, Thomas found giant, large-fruited wild fig trees and
"wild-growing, bitter, limes" fruiting abundantly, as well as an extensive
growth of frankincense trees at elevations of from 2,000 to 2,500 feet above
sea level. Search should be made in this region, between eastern
Hadhramaut and Oman, for the native home of the citron.
The gradual increase in use of the citron can be
traced in early literature. Theophrastus, writing at Babylon about
310 B.C., said the citron "was not eaten." Plutarch, writing
between 81 A.D. and 96 A.D., stated that "many substances which in the past
people would neither taste nor eat, are considered today as very
agreeable.…Shall we mention the cucumber, the melon, the Median apple, and
pepper?" (Tolkowsky, 1938, p. 91). By the second century the
epicurean Apicius Caelius could recommend the following dishes as being
exceptionally delicate: (1) the white inner part of the peel of citron made up
into a salad, and (2) small pieces of citron peel served with fish mixed with
herbs, vinegar, oil, and spices (Tolkowsky, 1938, p. 59). Before
long citrons had become a prized article of food in Rome, and in 301 A.D. the
records show that their sales prices were officially fixed by Diocletian at
values ranging from twelve to sixteen times the price of melons.
A method of candying citron peel was finally
discovered in the Mediterranean region. This involved the softening
and clearing of the peel, before in was candied, by fermentation in sea water
through the addition of a mixed culture of a yeast and a bacillus.
Candied citron peel ultimately supplanted almost completely the use of fresh
peel. However, as the fresh citron peel, like that of lemon and
orange, contains hesperidin (Penzig, 1887, p. 286), it is probable that it will
prove to be a good source of vitamin P because of its thick mesocarp, in which
the hesperidin is found. The peel should be tested for the making
of "citrin" (see also table 3-4 and below).
1a. Citrus medica var.
sarcodactylis (Noot.) Swing. Pl. Wilson. 2:141. 1914. Citrus
sarcodactylis Noot. Fleurs, Fruits, Feuill. Java 1:pl. 3. 1863.
Illus. Nooten, loc. cit., pl. 3; Swingle, in Bailey, Stand. Cycl.
Hort. 1:781. 1914.
Type.—Lacking
in Linnean Herbarium.
Distribution.—Widely cultivated in China, Japan, Indo-China, and
India.
Common name.—Fingered
citron.
Like the species except in the fruit,
which is split into a number of finger-like sections. Usually pulp
is lacking, or if present is very scanty.
The
fingered citron is well known and highly esteemed for its fragrance and beauty
in China and Japan, where it is called "Buddha's Hand Citron" (Fo Shou
kan in Chinese, Bushu-kan in Japanese). It is used by
Chinese and Japanese for perfuming rooms and clothing. It is also
grown as a dwarf plant, of which good fruiting specimens are highly prized for
ornamental purposes.
1b.
Citrus medica var. ethrog Engl. Die Nat. Pflanzenfam.
19a:338. 1931.
Type.—Europe
(Engler?). Herb. Bot. Mus.,
Berlin-Dahlem.
Distribution.—Western
Asia and Mediterranean countries; known only in
cultivation.
Common name.—Etrog
citron.
Fruits small, ellipsoid to fusiform,
rough, with a nipple ending in the persistent style with the persistent
stigma.
The "Etrog" of the Jews, used in the
Feast of Tabernacles, is not mentioned in the Bible. It probably
did not reach Palestine until after the time of Alexander the Great and was not
used by the Jews in fulfilling the prescriptions as given in Leviticus
23:40. Immanuel Löw (1924,p 286) stated that its use had been
recorded from the time of Alexander Jannaeus (104-78 B.C.).
On the basic of the detailed prescriptions
governing the character of the "Etrog" suitable for use in the Feast of
Tabernacles, it is highly probable that the present-day Etrog is a survival of
an ancient form of the citron, perhaps much like the original form of the
species from which the ordinary citron has been developed by selection.
2. Citrus limon (L.) Burm. f.
Fl. Ind. 173. 1768. Citrus medica var. limon L. Sp. Pl.
2:782. 1753; Limon vulgaris Mill. 1768; Citrus limonum
Risso, 1813; C. medica var. limonum Hook. f. Fl. Brit. Ind.
1:515. 1872. Illus. Loiseleur-Deslongchamps, Nouv. Duhamel
7:pl. 28. 1818; Risso & Poiteau, Hist. Nat. Orange. pl. 70 (col.).
1818-1822; Hayne, Arzn. Gew. 11:pl. 27. 1830; Bentley & Trimen, Med.
Pl. 1:pl. 54. 1880; Ford, Bot. Gaz. 104:288-305. 1942; and many
others.
Type.—Europe?
(L.?). Herb. Linn. in Linn. Soc.
London.
Distribution.—Southeastern Asia
(?); widely cultivated in all subtropical
countries.
Common name.—Lemon.
Small thorny trees, young leaves and flower buds
reddish; leaves pale green, long-ovate, pointed at the tip, margins serrate or
subserrate; petioles narrowly winged or margined, plainly articulated with the
leaf blade; flowers reddish-tinted in the bud; petals white above, purplish
below; stamens numerous, 20-40; ovary subcylindric or barrel-shaped, tapering
into the thick deciduous style; fruit oval with a broad, low apical papilla,
with 8-10 segments; peel yellow when ripe, rather thick, prominently
glandular-dotted; seeds small, ovoid, pointed, smooth, white within.
The origin of the lemon is a mystery.
Tanaka (1929b, p. 342) suggests that it was not introduced to China until
the Sung Dynasty (760-1297 A.D.), and it is still rare in India (according to
Biraghi, 1935). It was actively spread in the Mediterranean region
by the Arabs about 1000 to 1200 A.D. Like the sour orange, it
became widely and favorably known as a medicinal agent. According
to Glidden (1937, p. 382), the lemon was first described in detail by Ibn-Jami,
a physician at the court of Saladin (1171-1193 A.D.), in a treatise on the
medical uses of the lemon (now lost; parts, however, have been preserved in
quotations). The lemon was considered by Linnaeus to be a variety
of the citron (C. medica) and obviously closely related to
it. Probably the lemon should be considered as a satellite species
of the citron; possibly it may prove to be of hybrid origin, perhaps having the
citron and the lime for parent species. As is true with the
grapefruit, it is difficult to explain the origin of the lemon as a hybrid, as
it crosses readily with other species of Citrus and yet, when
self-pollinated, reproduces itself from seed with only small
variations. For this reason it is left here as a distinct, but
probably satellite, species of Citrus.23
The
glucoside hesperidin was found in fruits of the lemon tree by Pfeffer (1874,
p. 529). A comprehensive account of the biochemistry of the
glucosides, organic acids, sugars, vitamins, essential oils, and other
constituents found in the lemon is given in The Lemon Fruit by
Bartholomew and Sinclair (1951) and in U.S. Department of Agriculture
Agricultural Handbook 98 (1962).
The true
lemon is of recent introduction and is still very rare in southern China, as
was shown by Kwok Wa-shau, who in 1921 and 1922 made, under Swingle's
direction, for the Bureau of Plant Industry (now encompassed by the
Agricultural Research Service) a detailed survey of all the citrus fruits
cultivated or found growing wild in the vicinity of Canton. The
so-called white lemon, pak ning-mong in Cantonese, and the red lemon,
hung ning-mong, both of which grow commonly in southern China, are not
true lemons, but are hybrids, perhaps between the lemon and the mandarin orange
(see below).
The origin of the lemon
is still doubtful unless it proves to be a hybrid or sport of the so-called
lemon of India. The rough lemon, commonly used as a rootstock in
the United States, is apparently widely naturalized or possibly indigenous in
India and Pakistan, where it is called jambhiri. According to
Bonavia (1888-1890, p. 61), it was mentioned by the Emperor Baber in his
memoirs, composed in 1519 A.D. (Bonavia figured the fruit in pls. 131,
132). Hodgson (1937, p. 513) reported finding in India a form of
jambhiri "indistinguishable from our Florida rough lemon."
It grows in a seminaturalized state along the Mazoe River in Southern Rhodesia,
South Africa, where it is called the Mazoe lemon. The rough lemon
has been found to show a very high percentage of nucellar embryos, which would
indicate a hybrid origin. True lemons, on the contrary, show only a
small proportion (10 to 15 per cent) of nucellar embryony.
HYBRIDS OF THE LEMON
Lemonange ? [Citrus limon X C. sinensis ?].
The only hybrid of this supposed parentage that
has come into culture in the United States is the Meyer lemon, grown as a
potted plant in Peking, China, and introduced into this country in 1908 by the
late Frank N. Meyer, then agricultural explorer of the Foreign Plant
Introduction Service of the former Bureau of Plant Industry. It was
figured and described by McKee (1927, pp. 218-21, text figs. 48, 49); see also
Traub and Robinson (1937, p. 751). It is probably a hybrid between
the lemon and some other species of
Citrus.
Lemonimes [Citrus limon X C. aurantifolia].
These hybrids, made by safeguarded,
cross-pollinations, are intermediate between the two parent
species. The best known of these hybrids, the Perrine lemon (C.
aurantifolia 'Mexican' X C. limon 'Genoa'), is noteworthy in having
high resistance to lemon scab (caused by Elsinoë fawcettii) and also to
lime withertip (due to Gloeosporium limetticolum), the worst diseases
afflicting the two parent species. The Perrine lemon is described
and figured in Swingle, Robinson, and Savage (1931, p. 15, pl. 10), in McLennan
(1937, 1 text fig. and col. front cover), and in Traub and Robinson (1937, p.
781, fig. 15).
Lemandarins [Citrus limon X C. reticulata
?].
Probably the Otaheite orange, the hung
ning-mong, or "red lemon" of the Chinese of Canton, and the lemon called
pak ning-mong or "white lemon" are both hybrids of this character,
possibly complex hybrids. Tanaka has shown (1925, pp. 107-26, and
1933b, col. pl. 8) that the name Citrus limonia Osbeck belongs or
was assigned to one of these hybrids (not to the hung ning-mong, as he
supposes, but to the pak ning-mong), but such hybrid fruits propagated
artificially by man cannot be considered as true taxonomic species.
They arose doubtless as garden hybrids in southern China through crosses and
pollinations effected by insects between the lemon, possibly introduced by the
Arabs, and the native mandarin orange.
Rumphius (1741, vol. 2, p. 101, pl. 26, fig. 2) figured a lemon called Lemon
Martin, grown in the East Indian Archipelago by Europeans, that is almost
certainly a hybrid between a true lemon, probably brought in by the Portuguese,
and some other species of Citrus (possibly C. grandis or C.
hystrix); nevertheless it is called a lemon by the Europeans who live in the
East Indies and by other Europeans. The same thing seems to have
happened in southern China to the true lemon, probably transported there by the
Arabs and then degenerated by hybridization.
3. Citrus aurantifolia
(Christm.) Swing. Jour. Wash. Acad. Sci. 3:465. 1913. Limonia
aurantifolia Christm. in L., Pflanzensyst.…nach d. Houttuyn. 1:618.
1777; Limonellus sive Limon Nipis Rumph. Herb. Amboin.
2:107. 1741; Limonia acidissimia Houttuyn. (non L.) Natuurl. Hist.
2(2):444. 1774; Citrus lima Lun. 1814; C. acida Roxb. 1832;
C. spinosissima Meyer 1818; C. notissima Blanco, 1837; C.
limonellus, Hassk. 1848; C.medica var. acida Hook. f., Curtis'
Bot. Mag. 3 ser. 40:pl. 6745 (col.). 1884. Illus. Rumphius,
loc. cit. pl. 29; Merian, Metam. Insect., Surinam. 17, pl. 17 (col.).
1705; Hooker f. loc. cit. pl. 6745 (col.); Bonavia, Cult. Orang. Lem.
India Cey., Atl. pls. 228, 229. 1890; Hume, Cult. Citrus Fruits 19, 20, figs.
18, 20. 1926; Ochse, Fruits Fruitcult. Dutch East Ind. pl. 44 (col.). 1931;
European iconographies of Citrus usually have misleading illustrations
of this species!
Type.—East Indian
Archipelago, Amboyna, no type preserved; based on Rumphius' description and
plate and on Merian's plate.
Distribution.—East Indian Archipelago; widely grown in all subtropical
regions.
Common name.—Lime.
A small tree with rather irregular branches;
twigs with short, stiff, very sharp spines; leaves small, 5-7.5 cm long,
elliptic-ovate or oblong-ovate, obtusely pointed at the tip and rounded at the
base, margins crenulate, pale green, petioles narrowly winged, spathulate;
inflorescences axillary, short, lax racemes of 2-7 flowers (rarely single);
flowers small, white in the bud, calyx cupulate, 4-5 lobed; petals 4-5, 8-12 X
2.4-4 mm; stamens 20-25; ovary depressed, globose, with 9-12 segments, not
merging into the style but clearly set off from it; style soon deciduous;
stigma depressed, globose; fruits small, oval or subglobose, often with a small
apical papilla, greenish-yellow when ripe; peel very thin, prominently
glandular-dotted; seeds small, oval, white inside. (See Hodgson's
descriptions of limes in this work [excluding the Rangpur and Kusaie varieties, which
belong not to this species but probably to C. reticulata, or else are
hybrids of it with the lime].)
The lime is
apparently indigenous in the East Indian Archipelago. From there it
has spread by human help to the Asiatic mainland and to many other tropical or
subtropical regions of the world. It is a distinct species, not
closely related to any other species of Citrus. However,
limes hybridize freely with other species of Citrus and many such hybrids
are found in the East Indies. The Tahiti lime (see chap. 4, this work), which is widely grown in Florida and California, was
found by Bacchi (1940) to be in a triploid cytonomic state; Uphof (1931) had
already discovered that it produces no pollen grains or viable
ovules.
HYBRIDS OF THE LIME
Lemonimes
[Citrus aurantifolia X C. limon].
These hybrids, looking much like lemons, are
discussed above.
Limequats
[Citrus aurantifolia X Fortunella japonica, or F.
margarita].
These lime-like hybrids are discussed under Fortunella.
4. Citrus aurantium L. Sp.
Pl. 2:782. 1753. Aurantium acre Mill.Gard. Dict. ed. 8. 1768;
Citrus florida Salisb. 1796; C. vulgaris Risso, 1813; C.
bigarradia Loiseleur-Deslongchamps, 1818; C. bigaradia Risso &
Poiteau, 1818-1822; C. amara Link. 1831; C. karna Raf. 1838; C.
communis Le Maout. & Decaisn. 1868; C. aurantium var.
bigaradia Hook. f. 1875. Illus. Risso & Poiteau, Hist. Nat.
Orang. pl. 30 (col.). 1818-1822; Berg & Schmidt, Darst. Beschr. Off. Gew.
3:pl. 31. 1861; Bentley & Trimen, Med. Pl. 1:pl. 50. 1880;
Tschirch & Oesterle, Anat. Atl. Pharm. pls. 69, 70.
1898.
Type.—Europe ? (Linnaeus
?). Herb. Linn. in Linn. Soc.
London.
Distribution.—Southeastern
Asia; widely cultivated in all subtropical
regions.
Common name.—Sour or Seville
orange; bigarade.
A medium-sized tree up to 10
m high, with a rounded top; twigs angled when young, with single, slender
spines, often short, or stout spines up to 5-8 cm long on rapidly growing
shoots; leaves medium-sized, ovate, bluntly pointed at tip, broadly rounded to
cuneate at base; petioles 2-3 cm long, rather broadly winged, often 1.2-1.8 cm
wide at top, but sometimes narrower, 1 cm or less, narrowing rapidly to the
wingless base; flowers large, very fragrant with oil of neroli; 5-12 per cent
male (staminate only); fruits subglobose, usually slightly depressed at both
base and top, peel thick, with a rather rough surface, becoming brilliant orange
with a reddish tint at maturity; locules 10-12, filled with sharply acid pulp
and numerous seeds; fruit becoming hollow at center as it matures, and then
able to float in water. (See Hodgson's account of the cultivated
varieties of the sour orange in chap. 4 of this work).
This species was
introduced into the Mediterranean region from the East and for many centuries
was the only orange known to Europeans. During this long period of
culture it became very well known and much appreciated as a medicinal agent;
the fruits were used for flavoring and for marmalade, and the flowers for
perfumery. It was the orange of late medieval Europe.
Good high-flavored varieties of the sweet orange, C. sinensis, did not
reach Europe from southeastern Asia until the fifteenth century.
From that time on there was more or less confusion over the name of the sour
orange. The pharmacologists persisted in calling it C.
aurantium and the citrologists, then the botanists, called it C.
bigaradia. Many botanists considered both the sour and the sweet
orange as merely varieties of a single species. As a matter of
fact, the sour orange (C. aurantium) and the sweet orange (C.
sinensis) are very distinct botanical species, not merely cultivated
varieties of one species.
The chief
morphological differences are as follows: In the sour orange the petioles are
much more broadly winged than in the sweet, and the leaf blades are narrower and
more acutely pointed at the apex, and less rounded and more cuneate at the
base. Ruggieri has shown (1935) that the petioles of the sour
orange are much the longer, averaging 25.89 mm, whereas those of the sweet
orange average only 15.91 mm; in other words, the sour orange petioles average
63 per cent longer than those of the sweet orange. The fruits of
the sour orange are of a brighter orange color and have a rougher peel;
moreover, in the sour orange the oil glands are situated beneath minute
sunken areas in the peel, whereas in the sweet orange the tissue covering
the oil glands is often convex.
Uphof reported
(1932, pp. 133-35, fig. 4) that he found no male flowers on several cultivated
varieties of the sweet orange (C. sinensis), but that he did find from 5
to 12 per cent of male flowers on the sour seedling oranges of Florida (C.
aurantium). The sour orange, according to Uphof, "probably is
very close to the sweet orange but from the standpoint of the production of
male flowers constitutes a transition, so to speak, toward the lemons, limes
and citrons." Uphof found no male flowers on cultivated varieties
of the grapefruit (C. paradisi). Many trees of the Dancy
tangerine (C. reticulata) showed no male flowers (one small tree
overloaded with a very heavy bloom had one single male flower among the
thousands examined). Tangelos (hybrids of C. reticulata with
C. paradisi) also showed no male flowers.
In view of these facts, it is clear that the
occurrence in appreciable numbers of male flowers in C. aurantium
constitutes an important differential character separating this species from
C. sinensis.
The ethereal oil in the
leaves, flowers, and fruits of the sour orange is of very different odor than
that in the sweet (more agreeable and aromatic in the sour orange) and has a
different composition. Also the oil recovered from the petals of
the sour orange, neroli oil, finds a different use in perfumery and has a
higher value than that of sweet orange flowers. This oil,
considered by perfumery experts to be "indispensable to finer perfumery," is
said to owe its high value to small amounts (only 0.4 to 1.0 per cent) of "a
nitrogenous compound of exceeding fragrance," methyl anthranilate, NH2 · C6H4
· COOCH3 (see Gildemeister and Hoffmann, 1922,
pp. 93 and 96; and Finnemore, 1926, pp. 436-41). This remarkable
substance is not found in the oil extracted from the petals of the sweet orange
(Theulier, 1902). The pulp of the sour orange is intensely sour,
with a bitterish aftertaste, in contrast to the sweet, agreeable flavor of the
sweet orange. Furthermore, the peel of the sour orange, which is
official in the British Pharmacopœia (the peel of both the sour and the sweet
orange is official in the U.S. Pharmacopœia), contains three glucosides,
according to Tanret (1886, p. 518): (1) 4 to 30 parts per 1,000 of
isohesperidin (= naringin ?), having the same percentage composition and the
same rotary polarization as hesperidin but a bitter taste (hesperidin of the
sweet orange is tasteless) and a very different solubility; (2) 15 to 25 parts
per 1,000 of aurantamarin (to which, so Tanret asserts, most of the bitter
taste of the sour orange peel is due), differing slightly in composition and in
solubility from both naringin (?) and hesperidin; and (3) from a mere trace to
six parts per 1,000 of hesperidin. In the sweet orange only this
last glucoside, hesperidin, is found, but it is present in much larger
quantities than in the sour orange.24
The sour orange also shows physiological
differences from the sweet orange. It stands winter cold better
and has almost complete immunity to the foot rot, or mal di gomma, so
destructive to the sweet orange in some localities. However, the
sour orange is severely attacked by the scab fungus, Elsinoë fawcetti,
which does not attack the sweet orange.
Besides the morphological, chemical, and physiological differences mentioned
above, there are further anatomical difference discovered by Ruggieri (1935)
between the sour and the sweet orange in the separative layer of articulation
that lies between the petiole and the leaf blade, which may be summed up as
follows: (1) The pith in the lower articulation joint where the petiole joins
the twig is much more flattened from the top to the bottom in the sweet orange
than in the sour. In the sour orange the ratio of the pith to the
woody cylinder averages 1:1.4, measured in a horizontal direction, and 1:2.2,
in a vertical direction, whereas in the sweet orange the ratio of pith to woody
cylinder is 1:1.5 horizontally (about the same as in the sour orange) and 1:4.5
in the vertical (dorsiventral) direction, or only half as thick as the pith of
the sour orange petiole. (2) The cells of the interior layers of
the cortical parenchyma of the upper articulation joint where the petiole
joins the leaf blade are isodiametric and are 12 to 20.5 mu in diameter
in the sour orange but are 20 to 27 mu in diameter in the sweet orange,
or nearly one-half larger than in the sour orange. (3) The
pericycle fibers that form a more or less interrupted sheath around the woody
cylinder are strongly thickened in the sour orange but are little thickened, if
any, in the sweet orange.
B. Miyazaba, S.
Matsubara, and T. Kawaida (1928, p. 189, figs. 4, 5) have also found other
anatomical characters that separate the sour orange from the sweet.
Comparing the common sour orange (kaisei-to) of Japan with the
Washington navel orange (tento) in leaf structure, they found that the
sour orange leaf is thin (197 to 274 mu, the average of the ten
measurements being 243.38 mu), whereas the sweet orange leaf is about
11.1 per cent thicker (247 to 334 mu, the average of the ten measurements
being 270.33 mu). The two layers of palisade tissue are about
the same thickness in the two species, but the spongy tissue shows a wide
variation, being only 148 to 189 mu thick in the sour orange leaf (the
ten measurements averaging 167.28 mu) but ranging from 165 to 231
mu in the sweet orange leaf (the ten measurements averaging 197.51
mu), or nearly 17 per cent thicker. They also found that the
number of stomata is somewhat greater in the sweet orange epidermis than in
that of the sour orange, averaging 23.44 in the microscope field of vision for
the sweet orange and 20 for the sour orange, or 17 per cent more for the sweet
orange.
In view of this array of anatomical,
physiological and chemical differences between the sour and the sweet orange
it is obvious that they are distinct species even if the gross morphological
differences between them are small.
VARIETIES AND MUTATIONS OF THE SOUR ORANGE
Hodgson has given in this work an account of the forms of the sour orange known in the
United States, including the Bittersweet, with sweet fruits; the
Paraguay, with subacid fruits; and the Bergamot, grown in Italy
for the manufacture of bergamot oil from the fruits. These three
varieties are of unknown origin and opinions with respect to whether they are
mutations of the sour orange, like variety myrtifolia, or
hybrids. Experimental hybridization would probably settle the
question promptly.
The orangeries of Europe,
which came into vogue early in the fifteenth century and for two centuries were
among the most prized possessions of everyone who could afford one, were
filled with every form of citrus that could be found, among them many forms of
the sour orange. Tolkowsky (1938, p. 186) observed: "However, it was
the sixteenth and part of the seventeenth century…that gave citrus trees, both
in theory and practice, their final status as an element of the first
importance in the Italian garden." He also stated that the first
orange house or "orangery" in northern Europe, where citrus could not be grown
out of doors, was built by order of Charles VIII, king of France, in his own
château at Amboise after his famous expedition to Italy in 1494 and
1495. "The example set at Amboise was soon followed in other royal
castles and in those of many members of the nobility." The most
famous orangeries, according to Tolkowsky (pp. 200, 209), were those built for
Louis XIV at Fontainebleau and Versailles and finally, in 1674, the one for
Madame de Montespan at Clagny, where by 1675, so Madame de Sévigné reported to
her daughter, "there is a whole forest of orange-trees in huge boxes."
In 1818-1822, Risso and Poiteau in their famous
folio work, Histoire naturelle des orangers, published descriptions and
colored plates in natural size of 23 varieties of sour oranges called
bigarades. Of these, eleven had already been named, described, and
figured by Ferrari in his great work the Hesperides, published in
1646. A glance at these plates suffices to show what a wealth of
forms, both normal and teratological, were grown in Europe. Many of
these forms are still to be found in the citrus collections of the Old World,
and a few new forms have appeared in recent works on the European varieties of
citrus fruit trees (Riccobono, 1899, pp. 141-83; lnzenga, 1915, pp. 19-26;
Galli, 1928, pp. 10-13; and others). A striking new strain of the
sour orange, named the Oklawaha, was originated within the United
States. It has large fruits, 7.5 to 10 cm in diameter with a rind 6
to 10 mm thick, that are rich in pectin, making it an excellent marmalade
variety (see Swingle, Robinson, and Savage, 1932, pp. 9-10; and Traub and
Robinson, 1937, pp. 782-83).
The Trabut
variety of sour orange (named in honor of L. Trabut, who brought together at
Maison Carrée, Algeria, early in the twentieth century, a very fine collection
of cultivated varieties of citrus) is a European strain bearing normal
bitterish-sour fruits but differing from all other published descriptions or
figures of sour oranges in having very large, obdeltoid petioles, much broader
(2 to 3 cm) than those of typical sour orange leaves. It was
introduced into the United States by David Fairchild in 1925, and distributed
as F.P.I. No. 63550. It, like the Oklawaha variety just noted, is a
strain of sour orange very readily recognized by an easily seen morphological
character.
The Daidai is a cultivated
variety of the sour orange common in Japan that differs little from the
ordinary sour orange except in being dwarfish in habit, and having
blunt-tipped leaves and a persistent calyx that continues to grow as the fruit
develops, finally turning yellowish or orange-colored as the fruit
ripens. This variety shows almost complete immunity to citrus canker
in the Philippine Islands, where ordinary sour orange is very susceptible, as
Swingle observed in 1915. The Daidai is also known in China, where
it is grown in Kiangsu and Chekiang provinces for its flower buds, which are
used to mix with tea leaves. It is likewise grown in Canton,
especially in the famous Fa t’i flower gardens, as a potted plant.
Hu (1934, p. 47) described it as follows:
"After a growth of 5 or 6 years the Tai-tai [pronounced 'dai-dai'] tree
reaches a height of 1 m; its branches grow sparsely and are of spreading habit;
the leaves are elliptical or ovate, apex obtuse, base rounded; leaf blades are
very thick, petioles winged; leaves are broad and thick. Fruit is
compressed-globose, measures 5.4 X 6.4 cm; peel is of orange red
color. Fruit pulp is in 10 segments, membranes are thick and white;
fruit flesh is light yellow. Seeds ellipsoid, apex cuneate,
monoembryonic; cotyledons white in color; chalaza purple. Fruit
juice is very sour, so the fruits are not fit to eat in their raw
state. They [the Chinese orchardists] gather only the flower buds
of this species which they fire-dry to make a scenting material for tea
leaves."
Hu failed to mention that the Chinese
Tai-tai has enlarged, persistent calyx lobes, but a description drawn up by
Kwok Wa-shau, Swingle's former assistant working at Canton, China, from plants
introduced into Kwangtung Province (about 1915) from Soochow, Kiangsu Province,
reads: "The calyx is remarkably large, being strikingly different from that of
the other species of Citrus, bluish-green in color, the 5 sepals forming
a somewhat protruding cup…The rind is extremely thick and the pulp very
sour."
The Chinese do not use the Tai-tai as a
rootstock; instead they use the following sorts that grow to a larger
size:
The Vermillion globe sour orange
(chu-luan), which grows to a height of 5 meters and has long, narrow,
acutely pointed leaves. Hu (1934, p. 46) described it as having
compressed-globose fruits, 8 cm long and 9.5 cm wide, with many (30 to 40)
seeds. It is used as a rootstock for sweet oranges.
The Leather-head sour orange (p'i-t'ou
ch'êng), which has elliptical leaves, blunt at the tip, small
depressed-globose fruits, 4.4 cm long and 6 cm wide, with rough peel and with
numerous (about 20) seeds. It is used as a rootstock for sweet
oranges at Huangyen, Chekiang Province.
Tanaka, who studied the citrus fruit trees grown in the coastal regions of
China opposite Taiwan, reported (1932a, p. 29) that the sour orange
rootstocks used in Fuchow and Huangyen "are very different from our [Japanese]
Dai-dai." Probably the sour orange used as a rootstock at Huangyen
which Tanaka mentioned was the form called Leather-head sour orange by
Hu. Tanaka, who is familiar with the sour orange used as a
rootstock in the United States, stated further that "our Daidai…is the most
inferior grafting stock. On the other hand, the sour orange in the
United States and Italy is an excellent grafting stock, as well as that of
Chekiang…"
Benemerito (1938), like Kwok
(1922), found no typical sour orange growing in Kwangtung Province, only the
mutation Tai-tai, which has a large and persistent calyx. He listed
four other varieties that he assigned to Citrus aurantium, but three of
them, which have very narrowly winged petioles, are probably hybrids, whereas
the fourth, which has sometimes broadly winged and sometimes narrowly winged
petioles, was considered by Benemerito himself as probably a hybrid between the
sour orange and a mandarin. Some of these hybrids are worth testing
as rootstocks.
POSSIBLE ORIENTAL HYBRIDS OF THE SOUR ORANGE
Besides the four varieties from China and Japan,
discussed above, that seem to be mutations of C. aurantium, there are
other forms, probably accidental garden hybrids of unknown
parentage. The following are known from Taiwan and
Japan:
Hyonkan.
This form has large leaves (13 by 6.5 cm), acute
or acuminate at the tip, and small, subglobose (6 cm high, 7 cm wide),
thick-skinned fruits with a solid core. The calyx in the flowering
stage has slightly enlarged sepals; these, however, do not elongate as the
fruit grows. Hayata (1919, pp. 27, 28, text fig. 16) named it
Citrus daidai after a nomen nudum of Siebold printed in 1830, but
he stated: "It may be [a] hybrid between C. aurantium and C.
sinensis." It is reported as being cultivated in Taiwan and as
being much like a Japanese variety called Kwai-seikan ("very green [sour]
orange"). Certainly no evidence is adduced to show that this
variety merits a species name.
Nanshô
daidai.
This is a form found, very rarely,
growing semiwild in the forests of Taiwan in three localities, Nanchwang
(formerly Nanshô), Gaogan and Taitung districts. Tanaka
(1932a, pp. 20-21) stated that the "Nanshô daidai resembles the Daidai
[the Japanese form of] Citrus Aurantium in the shape of the fruits, but
the leaves are somewhat similar to the Naruto-mikan." He went on to
say that the fruits are round in shape with a slight nipple at the apex, have
an acid taste like the Yama-mikan, and have a thick rind like a citron.
The Nanshô daidai differs from the ordinary
Japanese Daidai, according to Tanaka (1926d, pp. 54-58), "in having much
longer leaves [blades about 12 by 5 cm], lanceolate, sharply and gently pointed
at the tips, thin, inodorous; petioles broadly winged; calyx glabrous, fruits
flattened, globose, peel with smaller oil glands, pulp delicate, very sour and
more deeply yellow." In this article the Nanshô daidai was
published in Japanese as Citrus taiwanica Tanaka and
Shimada. Tanaka (1926a, p. 345), a month later, published the
Nanshô daidai in English as a new species, Citrus taiwanica Tan., and
stated, "at present it seems to have become extremely scarce." It
is by no means clear that this is a good species; it is probably a hybrid of
C. aurantium with some other species of Citrus having long
leaves. Tanaka thought that the "very uniform" seedlings of this
species might "serve as stocks for commercially semitropic citrus
fruits." This very uniformity of the seedlings may be due to the
hybrid nature of the Nanshô daidai, as many Citrus hybrids yield
perfectly uniform seedlings from nucellar buds that replace the true embryo in
the seeds. In a later publication Tanaka (1933b, p. 172, fig.
138) included a halftone illustration of the type specimen showing flowering
twigs; another text figure (1933b, p. 136, fig. 112) illustrates the
depressed-globose, thick-skinned fruit, which shows 10
segments.
Yama-mikan ("Mountain
mandarin").
Concerning this form, T. and Y.
Tanaka (1932, p. 3) stated: "Yamamikan…is used as a rootstock in Hyûga Province
[in Japan], and it corresponds to the American sour stock in its vigor and
habit."
ACCURATELY SAFEGUARDED HYBRIDS OF CITRUS AURANTIUM
Apparently the only sour orange hybrid known
that was made by properly safeguarded cross-pollination is the following
bigeneric one:
Citradia [Citrus
aurantium X Poncirus trifoliata].
This beautifully intermediate trifoliolate hybrid has great vigor and hardiness
(see also under Poncirus).
4a. Citrus aurantium var. myrtifolia Ker-Gawl. [John
Bellenden] Bot. Reg. 4:text to pl. 346. 1818. Citrus myrtifolia
Raf. Sylva Tellur. Mant. 141. 1838; C. sinensis Pers. (non Osb.) Syn. Pl.
2:74. 1807. Illus. Ker-Gawler, loc. cit. pl. 346 (col.);
Risso & Poiteau, Hist. Nat. Orang. pl. 50 (col.). 1818-1822.
Type.—England (Ker-Gawl.).
Herb. ?
Distribution.—Known only in
cultivation: supposed to have been brought from China into the Mediterranean
regions in the seventeenth century.
Common
name.—Myrtle-leaf orange.
Twigs with very
short internodes, usually 1-2 cm long, often without spines; leaves small, about
1/2-1/3 the size of the normal foliage of the species; fruit small, often about
1/4-1/2 the size of that of the species.
This
plant is so strikingly different from the common orange that Charles Darwin
(1868, p. 100) considered it to be one of the most distinct species of
Citrus. He stated: "…I can adduce another case: the
myrtle-leaved orange is ranked by all authors as a variety, but is very
distinct in general aspect: in my father's greenhouse, during many years, it
rarely yielded any seed, but at last produced one; and a tree thus raised was
identical with the parent form."
The
myrtle-leaved variety of the sour orange probably arises as a mutation of the
common form of the sour orange. Swingle once found at Leesburg,
Florida, sprouts of this myrtle-leaved variety growing from the roots near the
base of the trunk of a very old sour orange rootstock on which a large sweet
orange had grown from a graft. Doubtless this old sour orange was
originally, like the many others found in the same grove, dug up from under
the live oak trees in a hardwood grove on rich soil (hammock) where sour orange
trees grew in a semiwild condition.
One
important horticultural race of sour orange, the chinotto of Italy, belongs to
this variety. It is grown for its small fruits, which are
candied.
THE TAXONOMIC PROBLEM OF CITRUS AURANTIUM AND ITS
FORMS
Citrus aurantium is unusual
and can almost be said to show mimicry of the sweet orange, C.
sinensis. It has required careful search to find morphological
differences to separate the two species; yet they show great differences in
their physiological requirements and limitations, in their graft compatibility
to other citrus fruit trees, in their resistance to disease, and especially in
the chemical composition of their essential oils, glucosides, etc.
If we admit that the two species are distinct in
spite of their mimicry, we then face the even more difficult problem of
distinguishing the similar-looking but physiologically diverse forms, whether
subspecies, varieties, or strains, found within the species. Two
forms of sour orange so similar as to look almost identical may have profoundly
different values as rootstocks for the lemon and perhaps for other cultivated
citrus fruit trees. This problem, already bafflingly complex, is
further complicated by the hybrids of C. aurantium, which make an
already bad taxonomic situation still worse.
5. Citrus sinensis (L.)
Osbeck, Reise Ostind. China 250. 1765. Citrus aurantium [var.]
sinensis L. Sp. Pl. 2:782. 1753; Aurantium sinensis Mill.
Gard. Dict. ed. 8. 1768; Citrus aurantium Lour. (non L.) Fl. Cochinch.
2:466. 1790; C. aurantium Risso (non L.), Ann. Mus. Hist. Nat.
Paris 20:181. 1813; C. aurantium [var.] vulgare Risso &
Poit. Hist. Nat. Orang. 33. 1818-1822; C. aurantium [var.] dulce
Hayne, Arzn. Gew. 11:pl. 28. 1830. Illus. Risso & Poiteau,
loc. cit. pls. 3, 4 (col.); Hayne, loc. cit. pl. 28 (col.);
Bentley & Trimen, Med. Pl. 1:pl. 51 (col.). 1880; and many
others.
Type.—Europe (Linnaeus), lost ?
(no authentic specimen in Linnean Herbarium, fide B. Daydon Jackson
[1912, p. 58]).
Distribution.—China,
Indo-China, possibly other southeastern Asiatic
regions.
Common name.—Sweet orange.
A medium-sized tree with a rounded top and
regular branches; twigs angled when young, usually with slender, somewhat
flexible, rather blunt spines in the axils of the leaves; leaves medium-sized,
pointed at the apex, rounded at the base; petioles narrowly winged, articulated
both with the twig at the base and with the leaf blade at the tip; flowers in
small racemes or singly in the axils of the leaves, medium-sized; calyx with 5
lobes; petals 5; stamens 20-25; ovary subglobose with 10-13 locules; style
slender, clearly delimited, soon falling; fruits subglobose, oval or flattened
globose; peel thin, tight, not bitter, central axis (pith) solid; seeds
cuneate-ovoid with rough-margined plane surfaces, white inside; embryos
usually numerous, varying greatly in size. (See Hodgson's
descriptions of the cultivated varieties of the sweet orange in this work.)
In 1914, in
discussing Chinese species of Citrus, Swingle stated (1914a, pp.
150-51): "The common sweet orange, often confounded with the sour or Seville
orange, is in fact quite distinct from the latter. The fruits of
the sweet orange have a solid core, never becoming hollow like that of the sour
orange; the petioles are narrowly winged in the sweet and broadly winged in the
sour orange; the leaves and flowers of the two species have a very distinct
odor. The two species show decided differences in their soil
requirements and in their susceptibility to the attacks of fungous
diseases. Many other minute but constant divergences are shown
between these two oranges in all their organs. These two plants,
then, superficially so similar, are in reality very unlike and should by no
means be united as varieties of one species." In the discussion
above under C. aurantium is has been shown that many anatomical
differences between the sweet and the sour orange have recently come to light,
differences that go far to show that these are in reality very distinct species
that happen to possess certain superficial resemblances concealing many
profound differences.
Citrus sinensis
is probably native to southeastern Asia, in northeastern India or, more
likely, in southern China and Indo-China. It has been so widely
cultivated for so long that it is no longer known in a truly wild condition.
Tolkowsky (1938) theorized that the sweet
orange first appeared in Roman gardens as early as the first century A.D.,
eventually disappearing in the barbaric invasions. The sweet orange
mentioned in literature of the fifteenth century appears to have reached Europe
over the Genoese trade route, followed by superior varieties brought around the
Cape of Good Hope by Portuguese navigators in the sixteenth century.
Before the advent of the sweet orange into the
Mediterranean regions, citrus fruits were esteemed there for medicinal use, but
the introduction of this delicious and beautiful fruit aroused an intense
interest in Citrus and led many people of means to build orangeries
wherein they grew all sorts of citrus fruits. Everyone in Europe
heard of these wonderful fruits. By the middle of the seventeenth
century (1646) Ferrari had written his Hesperides, a beautifully
illustrated account of the citrus fruits then known. Around this
was woven the story of Hercules, taken from Greek mythology, who was supposed
to have found oranges when he gathered the golden apples of the Hesperides.
The fruits of the
sweet orange contain a glucoside, hesperidin, which was discovered by Lebreton
in 1828 but was first recognized as a glucoside by A. Hilger (1876) and E.
Hoffmann (1876). It is allied to, but different from, the naringin
of the pummelo and the grapefruit, the aurantamarin of the sour orange, and the
ponciridin of trifoliate orange (see table 3-4). Much interest was aroused by the
discovery of "citrin," or vitamin P, related to hesperidin, in the juice and
peel of the lemon (and sweet orange) by Szent-Györgyi and his colleagues (see
Rusznyák and Szent-Györgyi, 1936; Armentano et al., and Szent-Györgyi,
1936; and Rosenberg, 1942). Further information on the composition
and biochemistry of the sweet orange may be found in Sinclair (1961) and the
U.S. Department of Agriculture's Handbook No. 98 (1962).
HYBRIDS OF CITRUS SINENSIS
Tangors [Citrus sinensis X C. reticulata].
These hybrids, which look much like the sweet
orange, bear orange-like fruits often of high quality. They are
discussed under C. reticulata.
Citranges [Citrus sinensis X Poncirus trifoliata].
See under Poncirus.
Citrangors [Citrus sinensis X (C. sinensis X Poncirus
trifoliata)].
See under Poncirus.
Citrangequats [(Citrus sinensis X Poncirus trifoliata) X
Fortunella japonica, or F. margarita].
See under Fortunella.
Citrangedins [(Citrus sinensis X Poncirus trifoliata) X
(Citrus reticulata var. austera ? X Fortunella sp.)].
See under Fortunella.
Citrangeremos [(Citrus sinensis X Poncirus trifoliata) X
Eremocitrus glauca].
See under Eremocitrus.
6. Citrus reticulata Blanco, Fl. Filip. 610. 1837. Citrus
nobilis Andrews (non Lour.), Bot. Repos. 9:pl. 608. 1809; C.
nobilis var. major Kerr, Bot. Reg. 3:pl. 211. 1817; C.
deliciosa Ten. Ind. Sem. Hort. Neap. [9]. 1840; C. nobilis var.
genuina Tan. Bot. Mag. Tokyo 26:204. 1912. Illus. Andrews,
loc. cit. pl. 608; Kerr, loc. cit. pl. 211; Du. Breuil, in Risso
& Poiteau, Hist. Nat. Orang. ed. 2. 49, pl. 29 bis.
1871-1872.
Type.—Wanting.
Substitute type: Philippines, Luzon (Merrill, Species Blancoanae, No.
402).
Distribution.—Philippines,
southeastern Asia; widely cultivated in all subtropical
regions.
Common name.—Mandarin
orange.
A small spiny tree with slender twigs;
leaves broadly or narrowly lanceolate; flowers arising singly or in small
clusters in the axils of the leaves; fruits depressed globose or subglobose,
with thin, loose peel easily separating from the segments, bright orange or
scarlet-orange when fully ripe; seeds small, pointed at one end, embryo
green. (See Hodgson's description of the cultivated varieties of
the mandarin orange in this work.)
In discussing the Chinese species of Citrus in Plantae
Wilsonianae (Swingle, 1914a, pp. 141-49), Swingle showed that the
true mandarin orange with its thin, loose peel is very different from Citrus
nobilis, which Loureiro described as having a "thick, succulent, sweet,
edible and irregularly tuberculate rind." As Swingle wrote at that
time, "this description is impossible to reconcile with the ordinary mandarin
oranges." It does describe the Cochin-Chinese orange that is called
King (or King of Siam) in the United States. This latter shows many
other important differences from the typical mandarin orange—differences so
great as to force the authors to the conclusion that it is a hybrid between a
mandarin orange and some other citrus fruit, possibly a sweet orange or a
pummelo, or even a pummelo X sweet orange hybrid. H. Brenier (1917, p. 214), in commenting on the confusion in the
names of Indo-Chinese citrus fruits, stated (in translation): "It may well be
that hybridization has occurred. It is indeed very probable that
the hybrid King of Siam (C. Aurantium X nobilis [i.e., C.
sinensis X C. reticulata]), an orange with high-colored pulp
mentioned in the Flore générale de l'Indo-Chine, is not the only hybrid that has originated in our peninsula."
In 1931, Matlack reported that the chromoplasts
in the peel of the King orange are very different from those of the mandarin
orange and similar to those of the sweet orange.25 Another
discovery, made by Nelson (1934), is of a fully methylated flavonol in the peel
of the tangerine orange. Nelson stated: "It seems to be the first
instance of a fully methylated flavonol occurring in nature." It is
doubtless analogous to hesperitin (derived from hesperidin of the sweet
orange), naringenin (derived from naringin of the grapefruit and the pummelo),
and ponciritin (found in flowers of Poncirus trifoliata by Hattori,
1936).
It thus became necessary to find
another botanical name for the mandarin orange. The oldest valid
published name for the mandarin type of orange is Citrus reticulata, so
named for the irregular network of soft, white fibers found between the loose
peel and the juice-filled segments. This name was published by
Blanco in his Flora de Filipinas in 1837. Because his type
specimens have disappeared, it was not entirely clear what plant Blanco had in
mind until Merrill (1918) published his study of Blanco's plants and issued
sets of herbarium specimens including substitute types for Blanco's new
species. Merrill's studies and substitute types show clearly that
Blanco's C. reticulata is, in fact, a mandarin orange. The
Blanco species name antedates by three years C. deliciosa Tenore, which
was based on the so-called Willowleaf mandarin. The Philippine C.
reticulata is fortunately not an extreme and highly specialized form of this
species, and hence is excellently fitted to be the type of this widespread and
variable species of Citrus. Probably a dozen or more
so-called species have been published for forms of mandarin oranges and their
hybrids and numerous varieties.
6a. Citrus reticulata var.
austera Swing. Jour. Wash. Acad. Sci. 32:25.
1942.
Type.—China, Kwangtung Province,
Ch'ao-chou (Groff, No. 233, 1918). Herb. Lingnan Univ., Canton,
China.
Distribution.—China: Swatow
region, Kwangtung Province.
Common
name.—Sour mandarin.
Differs from the
sweet mandarin orange in having smaller fruits with intensely acid
pulp. The type of this variety is the sour mandarin called sün
kat in Cantonese, propagated from seed in the Swatow region of Kwangtung,
where it is commonly used as a rootstock for grafting. G. W. Groff
in 1918, in a manuscript report covering his work on citrus done in China
under Swingle's supervision for the former Bureau of Plant Industry of the
U.S. Department of Agriculture, described the sün kat as
follows:
"Fruits slightly depressed-globose,
2.9-3.3 cm long, 3.3-3.6 cm diam., with smooth, loose peel about 4 mm thick,
capucine yellow (Ridgway, pl. 3) when ripe; oil glands small, round, far apart,
fragrant; segments 9, easily separated; segment walls thin, tender, white; core
6-8 mm diam., soft; pulp deep chrome yellow (Ridgway, pl. 3), composed of
small, short, pulp-vesicles, clinging together but irregularly arranged and
easily broken; juice reddish yellow, very sour; seeds about 9, rounded at one
end, pointed at the other, showing white parallel lines from base to tip;
leaves lanceolate-elliptical, blades 6.8 X 2.5 cm, rather acutely cuneate at
the base and narrowed to a blunt apex, with about 10 pairs of lateral veins;
petioles nearly wingless."
This variety is
widely grown about Swatow, China, where it is used as a rootstock upon which to
graft the Ponkan or mi-tong-kan ("honey pot orange") and other famous
varieties widely exported from Swatow.
Probably some of the other sour mandarins called kat by the Cantonese are
forms of this variety. Some of the so-called kat varieties
with large fruits, which as they ripen may become sweet enough to eat, are
probably hybrids between variety austera and the sweet mandarin, C.
reticulata, or the sweet orange, C. sinensis.
The so-called Rangpur lime (see chap. 4 this work) may upon further study prove to be a form of C.
reticulata var. austera or a dilute back-crossed hybrid of
it. It is probable that the sour mandarin has hybridized with
several other species of Citrus in China; hybridized with C.
ichangensis, it doubtless gave rise to the widely cultivated Yuzu, a sort of cold-resistant lemon-substitute widely cultivated
in Japan and northern China; hybridized with a kumquat (some species of
Fortunella), it gave rise to the well-known Calamondin, widely cultivated in China and the Philippines and
well known in citrus variety collections in the United States.
HYBRIDS OF CITRUS RETICULATA
The
following types of hybrids between mandarin oranges and other citrus fruit trees
are known:
Tangelos [Citrus reticulata X C. paradisi].
These hybrids, which look very much like oranges
and are often beautifully colored and of delicious flavor, were first made by
Swingle, using safeguarded cross-pollination technique. They were
described and illustrated by Webber and Swingle (1905, pp. 235-37, pls. 17-19);
by Webber (1907, pp. 336-37, pl. 21); by Swingle 1913e, pp. 87-89, pls.
on pp. 94, 95); and by Hume (1926, pp. 106-08). Additional tangelos
that had proved worthy of being named as horticultural varieties were described
and illustrated by Swingle, Robinson, and Savage (1931, pp. 1-13, pls. 1-7);
by Traub and Robinson (1937, pp. 766-67, figs. 6, 7); and by Hodgson in this
work (chap. 4 text, and fig. 4-64).
Similar hybrids
have originated in China, Indo-China, Japan, and other Far Eastern countries by
insect cross-pollination of mandarin oranges and pummelos growing in dooryard
groves of mixed varieties of citrus fruit trees. The Natsumikan or
Natsudaidai of Japan is almost certainly such an accidental hybrid; it cannot
be named as a new species of Citrus unless similar new species names are
coined for the many strikingly diverse tangelo hybrids that have been made in
the United States and with accurate knowledge of their parentage.
The name, Citrus natsudaidai Hayata, given by Hayata (1911-21, vol. 8, p.
29, fig. 19), cannot be accepted as a proper one for a chance hybrid known only
in cultivation.
Tangors [Citrus reticulata X C. sinensis].
These hybrids have been made by safeguarded
cross-pollination; among them is a promising tangelo-like fruit named the
Umatilla (see Swingle, Robinson, and Savage, 1931, pp. 11-12, pl.
8). The King orange of Indo-China, long grown in the United States,
is doubtless a tangor (see Brenier, 1917, p. 214; quoted above). The Temple orange of Florida is probably
another tangor (see Traub and Robinson, 1937, p. 775). There are
without doubt many tangors besides the King that are masquerading as oranges in
China, Japan, and other eastern Asiatic countries, where they have originated
as chance hybrids. As there are many more varieties of mandarin
oranges commonly grown in the Far East than in the United States, it is to be
expected that even more variation will be found in Oriental tangors.
The King tangor was made the type of a new
species which Loureiro (1790, vol. 1, p. 266) published as Citrus
nobilis. As Loureiro's herbarium specimens have almost all been
lost, this species was misunderstood by taxonomists, who considered it to be a
true mandarin orange, i.e., what is now called C. reticulata.
It is therefore necessary to abandon the name C. nobilis (see
discussion above).
Citrandarins
[Citrus reticulata X Poncirus trifoliata].
These trifoliolate hybrids are discussed under Poncirus.
Calamondin [Citrus reticulata var. austera ? X Fortunella
sp.].
See under Fortunella.
Yuzu [Citrus reticulata var. austera ? X C.
ichangensis].
See under Citrus
ichangensis.
Faustrimedin
[(Citrus reticulata 'Calamondin' X Fortunella sp.) X
Microcitrus australasica].
See under Fortunella.
7. Citrus grandis (L.)
Osbeck, Dagbok Ofwer Ostin. Resa 98. 1757. Citrusaurantium [var.]
grandis L. Sp. Pl. 2:783. 1753; C. aurantium [var.]
decumana L. Sp. Pl. ed. 2. 1101. 1763; C. decumana L. 1767;
Aurantium decumana Mill. 1768; Citrus pamplemos Risso, 1826; C.
maxima (Burm.) Merr. 1917. Illus. Loisleur-Deslongchamps, Nouv.
Duhamel 7:38, 42. 1818; Risso & Poiteau, Hist. Nat. Orang. pls. 62,
63. 1818-1822; Poiteau, Pomol. Franc. 2:pl. 344. 1846; Nooten, Fleurs,
Fruits, Feuill. Java 1:pl. 3. 1863; Tanaka, Kankitsu No Kenkyû (Citrus
Studies), pl. 3 (col.). 1933; Tillson & Bamford, Amer. Jour. Bot.
25:786, figs. 37-38. 1938; and many other plates (see Stapf, Index
Londinensis 2:223 [1930]).
Type.—China, Canton (Osbeck), in Herb. Mus. Stockholm (fide
Tanaka).
Distribution.—Southeastern
Asia, East Indian Archipelago; widely cultivated in all subtropical
regions.
Common name.—Pummelo.
A large, spiny, round-topped tree with angular
twigs, often pubescent; leaves large or very large, oval or elliptic-oval, with
a blunt point at the tip and a broadly rounded base, often subcordate and even
slightly overlapping the winged petiole; midrib and large veins often
pubescent; petioles broadly winged, and more or less cordate, usually
pubescent; flowers very large, borne singly or in axillary clusters or in
subterminal inflorescences; sepals and petals 5; stamens 20-25, with large
linear anthers; ovary globose, sharply delimited from the deciduous style, with
many segments; fruit large or very large, subglobose, oblate-spheroid or
subpyriform; seeds large, thick, wrinkled. (See Hodgson's
descriptions on horticultural varieties of the pummelo in this work.)
Citrus grandis
is one of the most distinct and most easily recognized species of
Citrus. It is separated from the other species of the genus
by a number of easily seen characters.
Its
thick, often pubescent, angular young twigs, with huge leaves borne on broadly
winged, more or less heart-shaped petioles, its very large flowers and giant
pale yellow fruits often the size of a child's head, make the species
impossible to mistake. The fruit usually has a thick peel and the
pulp-vesicles are much larger than those of other species of
Citrus. Instead of cohering with one another, they easily
fall apart. The membranes that enclose the segments, although
thin, are so strong that they can be peeled off the enclosed mass of
pulp-vesicles easily. If this is done carefully, the segment
remains intact in spite of having lost its covering membrane.
Besides the
morphological differences distinguishing it from the other species of
Citrus, the pummelo was discovered to have an important chemical
difference (later found to be shared by the grapefruit): it contains naringin,
a bitter glucoside related to the nearly tasteless hesperidin of the sweet
orange and to the bitter aurantamarin of the sour orange, but differing from
both. (Full information on naringin is given by Will, 1885 and
1887; see also Zoller, 1918, Poore, 1934; and table 3-4.) Certain Oriental pummelos have a slightly
higher content of vitamin C than oranges or grapefruit. Pummelos
are highly esteemed in both China and Thailand.
8. Citrus paradisi Macf. Hook.
Bot. Misc. 1:304. 1830; also Fl. Jamaica 1:131. 1837. Citrus
decumana var. racemosa Roem. Syn. Hesper. 1:67. 1846; C.
decumana var. patoniana Riccob. Boll. Ort. Bot. Palermo 7:211.
1908; C. maxima var. uvacarpa, Merr. & Lee, Amer. Jour. Bot.
11:383. 1924. Illus. Swingle, in Bailey, Stand. Cycl. Hort.
3:1392, pl. 50 and fig. 1744. 1915; Tanaka, Kankitsu No Kenkyû (Citrus
Studies), pl. 4 (col.). 1933; and many
others.
Type.—Wanting (fide
Tanaka, 1932e, p. 432).
Distribution.—West Indies; now cultivated in all subtropical
regions.
Common name.—Grapefruit.
A large, round-topped tree with dense foliage;
twigs angular when young, glabrous or nearly so; leaves larger than those of
the sweet orange, smaller than those of the pummelo, ovate, bluntly tipped and
broadly rounded at the base, glabrous or nearly so; petioles rather broadly
winged but not so broad as those of the pummelo, oblanceolate to obovate in
shape, the somewhat broadly rounded tip touching the very broadly rounded base
of the leaf blade; flowers large, borne singly or in small clusters in the
axils of the leaves; calyx 5-lobed; petals smaller than those of the pummelo,
often larger than those of the sweet orange; fruits larger than those of the
sweet orange but smaller than those of most pummelos; seeds smaller than those
of the pummelo, white, not yellowish, not ridged as are those of the
pummelo. (See Hodgson's description of grapefruit varieties in this work.)
The grapefruit
apparently originated in the West Indies. In spite of careful
search, it has not been found native in the Old World, where the parent species
is largely grown and where many horticultural varieties are known.
Another alternative is that the grapefruit may
be a hybrid of the pummelo with the sweet orange. The morphological
characters of the twigs, leaves, flowers, fruits, and seeds would suggest
this. The most important argument against such an origin of the
grapefruit is the absence of any breakup of self-pollinated grapefruit
seedlings into orange-like and pummelo-like forms. The fairly high
fertility of the grapefruit when pollinated by Citrus reticulata 'Dancy'
makes the absence of reversions in the self-pollinated seedlings of the
grapefruit hard to explain. On the other hand, the hybrids of C.
reticulata 'Dancy' with grapefruit show great variability; some of them are
grapefruit-like and others somewhat orange-like. In view of this
fact it seems best to retain the grapefruit, for the present at least, as an
independent but satellite species immediately following C. grandis, with
which it is so closely allied that many Citrus taxonomists consider it a
variety of that species. Patrick Browne (1756, 1789) was perhaps
the first to discuss the grapefruit, under the name "Forbidden Fruit" or
"Smaller Shaddock." Lunan (1814, pp. 171-73) was the first to use
the term grapefruit. He wrote: "There is a variety known by the
name of Grapefruit, on account of its resemblance in flavor to the
grape." Tanaka (1926b) gave the taxonomic history of the
grapefruit (in Japanese) and in the English summary a good technical description
of its botanical characters.
The fruit of the grapefruit contains a bitter glucoside,
naringin,26 first discovered in the pummelo. According to Zoller (1918,
p. 371), naringin is found in large quantities in the peel of the grapefruit
(from 0.66 to 0.80 grams per 100 grams of fruit). It occurs
without any hesperidin but seems able to give rise to a form of vitamin P,
according to Szent-Györgyi (1938). It is different from the
glycosides of hesperidin, found in the sweet orange, and aurantamarin, in the
sour orange.
It must be admitted that the
true nature of the grapefruit is still unknown. It is to be hoped
that the mystery of its origin can be settled by some of the newer methods now
used in taxonomic research.
HYBRIDS OF CITRUS PARADISI
Tangelos [Citrus paradisi X C. reticulata].
These orange-like fruits, some of them beautiful
in color and of delicious flavor, are discussed under Citrus reticulata.
9. Citrus indica Tan.
Studia Citrol. 2:164. 1928. Illus. Tanaka, Kankitsu No Kenkyû
(Citrus Studies), 94, fig. 80. 1933.
Type.—British India, Khasia, Churra, altitude 2,000-4,000 feet (Hooker f.
and Thomson, 15/8/50). Herb.
Kew.
Distribution.—Eastern Himalayan
region: "a really wild Citrus found in Nowgong District, Khasi Hills and
Manipur in Assam" (Tanaka, 1937, p. 235).
Common name.—Indian wild orange.
The
original description, translated into English, reads as follows: "Branches
terete, spiny, glabrous; leaves oblong or lanceolate, thick, sub-entire,
attenuate at the apex, acute at the base, veins curved, minute, indistinctly
reticulate; petioles articulated, linear, sulcate; flowers not seen; fruits
small, broadly obovoid or sub-pyriform, solitary on terminal twigs, pedicels
very short; calyx 'hypocrateriform,' lobes triangular, acute; cortex thin,
segments few (?), pulp vesicular, vesicles fusiform; seeds large,
suborbicular, smooth, mono-embryonic." In his discussion Tanaka
stated that this species "has leaves resembling those of C. sinensis, but
[the] small, fig-shaped fruit containing extremely large seeds is entirely
different from any [other] Citrus."
In
the Japanese account of C. indica, Tanaka gave measurements of various
organs and the details not treated in his Latin diagnosis. His
detailed description in Japanese (1928a, pp. 160-62), as translated by S.
Katsura, reads: "Twigs round in cross-section, rather stout, somewhat zigzag
('wild-goose-procession form'), surface light green, glabrous, not wrinkled;
thorns strong, arising at a broad angle, sharp-pointed; leaves oblong or
lanceolate, pointed at both ends, apex somewhat caudate, the extreme point
sharp or slightly emarginate, the upper side green with indistinct veins, the
underside light-colored with fine curved veins, without convex oil glands,
margin subentire; petioles short, with narrow wings articulated [with the leaf
blade]; leaves near the top of the branches measure 12.2 X 4.1 cm, petioles 0.8
X 0.2 cm; flowers indistinct [only calyx seen]; fruits terminal, borne singly
on the ends of branchlets, pedicels extremely short and strong, articulated
with the calyx; calyx funnel-shaped, sepals triangular, acute, thick, glabrous;
young fruits depressed at apex, slightly attenuate at the base, short,
pyriform, about 2 cm diam., peel very thin [red, fide Tanaka, 1937],
segments few; pulp-vesicles spindle-shaped, stalked, very delicate; pulp very
slimy [fide Tanaka, 1937]; seeds very few, very large, almost round [in
outline], flattened, monoembryonic, 11 X 9 X 3.5 mm."
In discussing specimens collected at Khasia by
Hooker and Thomson in August, 1850, and preserved in Kew Herb. (Tan. Ident. No.
1851), Herb. Bot. Mus., Berlin-Dahlem (Tan. Ident. No. D-1282), and Rijks Herb.,
Leiden (Tan. Ident. No. R-1016), Tanaka stated that they "have branches with
numerous spines and luxuriant growth of leaves. The calyx persists
after the flower has fallen. Flowers borne single in the axils of
the leaves, with extremely short pedicels. Calyx tubular (fistulose
?) articulated with the pedicels. Ovaries vase-shaped and scarred
where the style has fallen off, resembling ovaries of Lavanga
[=Luvunga]. Disk noticeable, below the ovary." In
his remarks in Japanese, Tanaka stated: "The most outstanding point [of this
new species] is the morphological aspect of the flower parts. The
Leiden specimen has sepals pointing [out] like claws with oil cells on the
upper side resembling those of Chalcas [=Murraya], the flowers are
evidently borne singly.
"It resembles
Metacitrus [Citrus junos, C. ichangensis, and the loose-skinned
oranges, C. reticulata and its cultivated varieties and hybrids] in its
short pedicels, the thin skin of the fruits and the large seeds like C.
yuzu and C. ichangensis but in its depressed shape it resembles Pleiospermium [one of the Primitive Citrus Fruit Trees].
"In the shape of the articulation of the calyx
and pedicels, it resembles Citron and also in the round branches in cross
section, but its leaves are very different because they have no dentations at
all. The leaves show analogies to those of the sweet orange ?
('amodaidai') group in shape, but are longer and have far more indistinct fine,
parallel and curved veins. The form of the large leaves borne
scattered on strong shoots resembles that of leaves of the genus
Paramignya, in short it is still undetermined to which group this species
belongs."
In a later publication, however,
Tanaka (1937) stated: "It is really a wild Citrus found in Nowgong
District, Khasi Hills and Manipur in Assam. It belongs correctly to
Metacitrus, as was predicted by the author, having small sized fruit with
thin reddish rind and very slimy pulp, much similar to that of Citrus
depressa Hayata. Perhaps of value as a rootstock."
This remarkable species has several unusual
characters which indicate that it may be a truly indigenous wild
species. There is always the chance, however, that C. indica
is in fact a hybrid of the remarkable, truly wild Citrus species of
northeastern India, C. latipes, which probably has very large seeds like
its close relative, C. ichangensis, of central and southwestern
China. If so, hybrids between Indian C. latipes with some
species of the subgenus Citrus could be expected to have very large seeds
like the Ichang lemon, which very probably is a hybrid of C.
ichangensis and C. grandis.
10. Citrus tachibana (Mak.) Tan. Bul. Sci. Fak. Terk. Kjushu
Univ. 2:52. 1926. Citrus aurantium var. tachibana Mak.
Jour. Jap. Hort. Soc. No. 75:2, pl. 1896; C. nobilis var.
spontanea Tokutaro Ito, Jour. Coll. Sci. Univ. Tokyo 12:361. 1900;
C. aurantium subsp. nobilis var. tachibana Mak. Bot. Mag.
Tokyo 15:167. 1901. Illus. Makino, loc. cit. 1896; fig. 3-46 this work.
Type.—Lost. Paratype: Japan, Tosa (T.
Makino). Herb. Hokkaido Imp. Univ. (Tanaka ldent. label
Sa-2).
Distribution.—Southern Japan:
Yamaguchi and Kochi prefectures, south to Kagoshima Prefecture; Ryukyu Islands;
Taiwan.
Common name.—Tachibana
orange.
A description of this species by T.
Makino, as translated by S. Katsura, reads: "Tree stands over 10
feet. Branches and leaves grow thickly. Strongly
resistant to frost or snow. Fruit somewhat flattened, 2-3 cm
lateral diam. Skin smooth, oil glands scattered beneath the
skin. Segment cases 6-7; juice bitter and almost
inedible. Seeds 1-2 in a segment, and rather large in
size. Fruits at first green but turning yellow in the late
autumn. The flavor of the skin resembles that of Yuzu. Flowers the same as other Citrus plants in time
of blooming, shape and color. In the rainy season of summer the
petals begin to turn white. Needle-like thorns are found on the
branches. Petioles not winged."
A
description of the paratype in the herbarium of Hokkaido Imperial University
made by Tanaka reads as follows: "Branches small and slender; thorns about 3 mm
long, acute. Leaves long, ovate-elliptical, subcoriaceous, broadly
acuminate, obtuse and incised at the tip, somewhat broad and convex at the
base, indistinctly dentate at the margin, midrib slender, straight and distinct
beneath, veins almost indistinct, oil glands indistinct; petiole short, small,
with linear wings which seem to be on the verge of degeneration.
Flowers axillary, solitary, small. Pedicels 2 mm long, slender,
glabrous; scales at the base triangular, ciliate at the margin.
Calyx 3 mm in diameter; sepals somewhat recurved outward, densely ciliate at
the margin. Disk large, ring-form, subcarnose, depressed at the
apex. Ovary almost globular, attenuate at the base, about 2 X 2 mm
in size."
Tanaka (1922, pp. 246-48) published
in English a very interesting account of the tachibana orange, about which
cluster many of the oldest Japanese traditions, showing that the Japanese
people have known this fruit since the very beginning of their
history. As this species is not edible, it would doubtless not
have attracted so much reverent attention had it not been the only citrus fruit
known to the early Japanese.
The widespread
occurrence of the tachibana, from southern Taiwan to the southwestern province
of the main island of Japan, makes it very probable that it is in fact a wild
species that has persisted since prehistoric times. Tanaka
(1931a) published in Japanese a detailed account of the occurrence of
C. tachibana in a wild condition in southern Japan, on the Ryukyu
Islands, and also of its discovery in southern Taiwan where it grows at much
higher altitudes than in Japan but where the temperatures are much the same as
at sea level along the northern limits of its range.
This species is very similar in many of its
characters to the mandarin orange, C. reticulata. The great
antiquity of the tachibana in Japan seems to preclude its being a hybrid or
"chance seedling" of recent origin. It probably should be
considered as a satellite species of C. reticulata and somewhat closely
related to it.
POSSIBLE HYBRIDS OF CITRUS
TACHIBANA
Shekwasha (also spelled
Sheequasha, Shiikuwasha, Sekwasa, or even Seequassia).
This plant was described by Hayata (1919, p. 16)
as follows (in translation): "Twigs very green, applanate-triangular, obtusely
bent at the node, sometimes with spines 1-1.5 cm long in the axils of the
leaves. Leaves ovate-oblong, 8-9 cm long, 3.5-4 cm wide, slightly
acuminate or broadly obtuse and retuse at the apex, broadly and obtusely
triangular at the base, margins subentire, slightly crenulate; petioles 8 mm
long, very narrowly winged. Fruits terminal, with very short
peduncles, depressed-globose, 23 mm long, 4.5 cm wide, rounded in outline or
sometimes slightly lobed, deeply sunken at the apex and slightly sunken at the
base, with 7-9 locules, pericarp very thin, 1 mm thick, luteo-flavescent,
rather smooth, flesh acid. Seeds 1 cm long, 6 mm wide, apex acute,
rostrate tip obtuse; embryo pale greenish."
Tanaka (1927c, p. 31) added a few points in his short description:
"Bushy tree about 4 m high. Leaves small, broad-elliptic, obtuse,
petiole short, almost wingless. Fruit small, oblate, ends concave,
very smooth, oil cells fine, reddish orange. Rind thin,
turgid. Segments many. Center hollow. Pulp
deep-colored, sweetish and rich in pectin, vesicles finely netted.
Seeds very plump, finely striated, teguman [sic] at chalaza rosy,
polyembryo green." He figured the Shekwasha (1933b and
1926c, pp. 190-93, figs. 1, 2) in comparison with Citrus
tachibana, from which it differs strikingly in having larger oil glands
distinctly sunken in the peel, also fewer locules that separate from each other
and from the peel in the ripe fruit. Also Tanaka (1927b, p.
199) contrasted the Shekwasha with the Koji orange of Japan (which he called
"Citrus leiocarpa Hort. nov."), from which it differs in never occurring
north of Sanbok (in the Ryukyu Islands), and "by having pointed leaves, reddish
rind, finely round-netted pectiniferous pulp-vesicles, large seeds of brownish
coat and dark brown chalaza part of tegumen."
The Shekwasha is found, according to Tanaka (1927c, p. 31), in a wild or
semiwild state in the Ryukyu (Luchu) Archipelago south of Sanbok and in
Taiwan. Swingle brought back to the United States in 1927 a plant
of the Shekwasha given him by the Tanaka Citrus Experiment Station at
Tanushimaru, Kyushu, Japan, to which was assigned the Foreign Seed Introduction
number 71180 and C.P.B. number 936. In this country the fruits of
this variety when fully ripe are bright orange-colored and the less mature
fruits are orange-buff or capucine yellow; the pulp is near primuline yellow
(Ridgway, 1912, pl. 16). The seeds are pointed-obovoid and
somewhat compressed, 11 to 13 by 7 to 8 by 5.5 to 6 mm, with a very finely
striate but nevertheless remarkably smooth and even testa for a Citrus;
the chalazal cap is reddish-brown (Ridgeway, pl. 27); the embryo is pale
greenish. Tanaka reported (1926c, p. 192) that the pulp of
the Shekwasha contains much pectin.
The
Shekwasha grows so vigorously in the peculiar sandy-loam and porous-limestone
soils of extreme southeastern Florida that it should be tested there as a
rootstock. The fruits are poor-flavored.
Hayata (1919, p. 16) named the Shekwasha as a
valid species of Citrus (C. depressa); in the same paper, however,
he named as "new species" of Citrus several other forms that, according
to his own statement, are probably hybrids. The Shekwasha has just
the characters that would result from the hybridizing of the native C.
tachibana of Taiwan and the Ryukyu Islands with some form of the mandarin
orange (C. reticulata) introduced from the Chinese mainland.
Subgenus Papeda
Common
name.—Papedas.
Pulp-vesicles with very
numerous droplets of acrid oil; petioles always large and broadly winged;
flowers small, usually less than 2 cm diam.; stamens free, or, if cohering in
bundles, then flowers larger (1.5-3 cm diam.) and petioles very long, 1 3/4-3
times longer than broad.
This subgenus
comprises a number of truly wild species of Citrus, many of them still to
be found growing in primeval forests of the Monsoon region. There
are two sections in the subgenus Papeda, the section Papeda,
comprising the typical species, and the section Papedocitrus, which is
intermediate in character between the subgenus Citrus and the section
Papeda, having flowers much like those of the subgenus Citrus and
leaves like those of the section Papeda. The fruits are
almost never eaten, but the acrid juice of the fruits of the species belonging
to the section Papeda is widely used by primitive peoples, especially
Malays, Melanesians, and Polynesians, as a hair wash.
As noted above in the discussion of the subgenus Citrus, the
species of Citrus belonging to the subgenus Papeda have been shown
by Tillson (1938, pp. 21, 30; also Tillson and Bamford, 1938, pp. 788, 790) to
possess a decidedly simpler type of vascular anatomy of the flowers than do the
species of the subgenus Citrus. In this important character
the subgenus Papeda agrees with the other five genera (other than
Citrus) included in the True Citrus Fruit Tree group. This
fact will doubtless prove important in studying the course of evolution of the
commonly cultivated species of Citrus, all of which belong to the
subgenus Citrus. This makes it very desirable to learn much
more than we now know regarding the species of papedas in their relationship to
the subgenus Citrus, on the one hand, and to the genera Clymenia,
Microcitrus, Eremocitrus, Poncirus, and Fortunella,
on the other. Tillson's discovery of a primitive character in the
papedas serves to emphasize how very different they are from the true oranges
of the subgenus Citrus.
The species
which belong to the section Papeda have small flowers, 1.2 to 1.7 (rarely
2) cm in diameter, with free stamens not cohering in groups. The
pulp-vesicles of all the species contain numerous globules of very acrid oil and
are sometimes attached to the radial locule walls for one-half to three-fourths
the distance from the dorsal wall to the center of the fruit.
To facilitate an understanding of the subgenus
Papeda, the detailed key was prepared to give a perspective view, as it were, of the
taxonomic characters of the species that could not easily be obtained from
independent descriptions, no matter how detailed they might be.
Subgenus Papeda Section Papedocitrus
Flowers large (1.5-3 cm broad when open); stamens
at first coherent nearly to the tips, but separating later into a few bundles,
connate only at the base; winged petioles long-elliptical with more or less
parallel sides; seeds crowded, often very short, very thick, and angular.
The section Papedocitrus comprises two
species growing in southwestern China, northeastern India, and northern Burma,
far to the north of the range of the species that constitute the section
Papeda. Section Papedocitrus is intermediate in its
flower characters between the section Papeda and the subgenus
Citrus, but the pulp-vesicles contain numerous droplets of acrid oil and
the winged petioles often equal the area of the leaf blade, just as in the
section Papeda.
The fruits of the
species of this section do not seem to be used by primitive peoples for washing
the hair, but it is believed by some that in China the fruits of C.
ichangensis were anciently, and perhaps still are, used in
medicines. Striking hybrids have arisen in China between this
species and species of the subgenus Citrus. One of these
hybrids, the Ichang lemon, is cultivated on a small scale by the
Chinese. The Yuzu, which shows many points of similarity with C.
ichangensis, is doubtless another of these hybrids; it has been known in
China since ancient times and is widely grown both in northern China and in
Japan.
11. Citrus
ichangensis Swing. Jour. Agr. Res. 1:4 (excluding fruit). 1913.
Illus. Swingle, loc. cit. pl. 1, figs. 1, 4-7 (excluding figs. 2,
3 of fruit); fig. 3-47 this work.
Type.—China, Hupeh Province, Hsing shan district (E. H. Wilson).
Herb. Arnold Arbor., Harv. Univ.,
Cambridge.
Distribution.—West-central
and southwestern China: Hupeh, Szechwan, Kweichow, and Yunnan
provinces.
Common name.—Ichang
papeda.
A spiny shrub or small tree usually
5-15 ft. [1.5-4.6 m] high; twigs angular when young, with stout, sharp spines,
1.5-2.5 cm long, often reduced or lacking on flowering twigs; leaves narrow,
4-6 times longer than wide, mostly 8-11.5 X 1.8-3 cm; petioles very large,
broadly winged, obovate or oblong-spatulate, evenly rounded at the tip and
narrowed abruptly at the base, usually 3.5-6 X 2-3 cm; leaf blade
ovate-acuminate, often more or less caudate, emarginate at the tip and evenly
rounded or bluntly pointed at the base, usually 3-6 X 1.8-3 cm, often not
equaling the winged petiole in area; flowers 2.5-3 cm diam., 5-merous; sepals
thick, 3 mm long and 3 mm wide, with minutely ciliate margins; petals oblong,
1.5-2 X 0.5-0.8 cm, white; stamens 20, at first all connate to the tips,
finally breaking up into several bundles, about 10 mm long; pistil about 10 mm
long; style very short, caducous; stigma nearly as large as the ovary; ovary
with 7-9 locules, ovules numerous in each locule; fruits small, glabrous, 3-4
cm diam. in dried specimens (probably 3.5-5 cm when fresh), peel rough, 2-4 mm
thick in dried specimens; seeds large, very thick, 12-18 X 12-18 X 7-9 mm in
dried material, very blunt at both ends, with a chalazal cap about 12 X 5 mm,
apparently monoembryonic.
This species differs
from its congeners in having large, very thick seeds and slender leaves 4-6
times longer than broad, with very large, winged petioles often as large or
larger than the blade. It differs from C. hystrix in having
oblong, rather than triangular, winged petioles and much larger flowers with
connate stamens. (See fig. 3-47.)
This remarkable
plant,27 which grows in a truly wild state in central and
southwestern China, is doubtless the most cold-resistant of all the evergreen
species in the orange subfamily. It differs greatly from the other
species in the subgenus Papeda in having large flowers, partly connate
stamens, and large, very thick seeds, which are nevertheless
monoembryonic. The leaves also differ from those of all other
species of Papeda in having extremely long but rather narrow winged
petioles, often exceeding in area the slender, pointed, lanceolate leaf
blades.
In 1926, Swingle brought to this
country a healthy young seedling of this plant from Hupeh Province, grown for
him at Nanking, China, by C. C. Hu. As C. ichangensis is the
hardiest known evergreen species of Citrus, it is of great interest for
use in breeding cold-resistant hybrids, such as the Ichang lemon and
Yuzu.
HYBRIDS OF CITRUS ICHANGENSIS
Ichandarins [Citrus ichangensis
X C. reticulata cv. satsuma group].
An ichandarin was produced by the citrus experts of the former Bureau of Plant
Industry (now encompassed by the Agricultural Research Service) by applying the
pollen of the Ichang papeda to properly prepared flowers of the satsuma
orange. Naturally all chances of foreign pollen reaching the
crossed flowers were excluded, and of course the stamens of the satsuma flowers
had been cut away before any pollen had matured.
The leaf blades of this hybrid are broadly
elliptical, 6 to 8.5 by 2 to 4 cm, slightly acuminate at the tip and cuneate at
the base (angle 70° to 85°), margins minutely serrulate, dentate above the
middle. The leaf blades differ only slightly from those of the
Yuzu, a hybrid in which the leaf blades are more acuminate at the tip and more
broadly rounded at the base. The winged petioles are obovate,
oblanceolate, or elliptical, broadly rounded at the tip and gradually narrowed
into the base when the petiole is merely margined. The petioles are
so much like those of the Yuzu that it is often impossible to distinguish
between them with certainty. This hybrid is of interest chiefly in
that it throws light on the origin of the Yuzu, a well-known Chinese and
Japanese citrus fruit tree.
It is very
probable that if some strain of C. reticulata other than the satsuma, for
example the tangerine or the Willowleaf mandarin, had been used as one of the
parents the leaf blade would have been more acuminate at the tip and more
broadly rounded at the base, i.e., like the leaf blade of the Yuzu, which is
doubtless such a hybrid.
Yuzu [Citrus ichangensis X C. reticulata var.
austere ?].
Illus. Takahashi,
Kankitsu Saibai 112, fig. 31. 1912; Makino, Shokubutsu Dzukan 1064, fig. 2022.
1917; Tanaka, Jour. Hered. 13:pl. facing p. 243. 1922; Hu, Nogyo Oyobi
Engei 5:1637-1638, figs. 34-37. 1930; idem, Calif. Citrog.
16:544, pl. 2, fig. 15. 1931; idem, Jour. Agr. Ass. China Nos.
126-127:58, fig. 15. 1934; Murakoshi, Nai-gai Shokubutsu Genshoku
Dai-zukan 5:118, pl. 24, fig. 251 (col.). 1935.
The Yuzu is a medium-sized spiny tree; leaves
lanceolate-acuminate, with rounded bases but with pointed, usually acuminate
tips, slightly crenulate-margined toward the tips, leaf blades 5-7 X 2.5 X 3.5
cm; winged petioles obovate, 18-30 X 6-15 mm, with entire or very faintly
crenulate margins; fruits depressed-globose, usually with 10 locules, 5-7 cm
diam., 4.5-5.5 cm high, with a rough, bumpy peel, greenish in color when ripe;
pulp very acid, and somewhat bitterish; seeds plump, about 12-14 X 7-8 X 6-7
mm.
This well-known citrus fruit tree of
China and Japan is doubtless another ichandarin resulting from an accidental
cross-pollination of some cultivated variety of the mandarin orange by the
Ichang papeda, probably accomplished by some insect. The Yuzu is
unlike either parent species in many important taxonomic characters; however,
there has been little opportunity to learn its characters or experiment with it
since one parent, C. ichangensis, is a wild species (apparently never
cultivated in China) that was not discovered until 1913. The hybrid
nature of the Yuzu was not known even in the Orient, where C. ichangensis
is a botanical curiosity. As has been described above, hybrids that show astonishing similarity to the
Yuzu have now been produced in this country between the Ichang papeda and the
satsuma orange (a form of C. reticulata).
The Yuzu is sparingly cultivated in
north-central China, in Kiangsu, Chekiang, Hupeh, and Kansu provinces, and in
the plateau regions of southwestern China and as far south as Yunnan
Province. It is more commonly grown in Japan, both for its acid
fruits, which are used as a substitute for lemons or limes, and as a rootstock
for the satsuma and other cultivated varieties of citrus fruits.
Meyer, in October, 1914, found this hardy citrus
fruit tree in northwestern China, in the latitude of Atlanta, Georgia, at
Hsi-Chi village, near Siku (Lat. 33° 44' N., Long. 104° 30' E.), in the
southern part of Kansu Province. It was growing at an altitude of
610 to 1,372 meters (2,000 to 4,500 ft.) along with walnuts, persimmons,
pomegranates, and the Trachycarpus palm. Meyer (1918) described the
fruit as follows: "The fruits were loose-skinned, round flattened, the size of
mandarin oranges, color of rind light yellow; rind full of oil glands, smelling
like a fine lemon; segments separating easily; fairly juicy and of an
agreeable sharp sour taste; contains plenty of large seeds."
Meyer's photograph, taken at His-Chi, Kansu Province, was published by Tanaka
(1922, pl. facing p. 243).
Two strains of this
hybrid grown in north-central China were described by Hu (1934, pp.
47-48). They are: (1) hsiang ch'êng (aromatic ch'êng), a
name current in Chekiang and Kiangsu provinces; and (2) Lo han ch'êng
(Buddhist disciple Ch'êng), a name used in T'ang-ch'i, Chekiang
Province. Hu (1930) has shown the first in his figures 34 and 35
and the second in figures 36 and 37.
Tanaka
has directed attention (1933b) to certain Chinese records which seem to
prove that this plant was known and cultivated in ancient China under the name
yu. This name is still used for it in Japan but not in China,
where the name is now applied to the pummelo (C. grandis), a very
different species having much larger fruits with agreeably flavored sweet
pulp. The Yuzu is now called ch'êng tzu in China and was so
called as early as 1108 A.D. by Tang Shên-wei in his great illustrated herbal,
the Chêng lei pên ts'ao, published in that year.
In the "Spring and Summer Annals" of Lü Pu-wei,
who died in 237 B.C., the yu of that epoch was described as follows (as
translated by Michael J. Hagerty): "Some are sweet and some are
sour. The sour are called hu kan or Barbarian
sweet. At present the common people sometimes speak of the
ch'êng as yu but this is wrong." This last sentence
proves, as noted by Hagerty, that there were already in the third century B.C.
two different fruits called yu. It seems probable that the
Japanese name yuzu, which corresponds to yu tzu in the Chinese
spoken language, has been kept with its ancient meaning in Japan, but that the
meaning has for many centuries been lost in China.
The Yuzu was named Citrus junos (as a good
species) by Tanaka, but, as has been shown above, it is very probably a hybrid
of the Ichang papeda and some Chinese cultivated variety of the mandarin orange
and therefore cannot be recognized as a good botanical species.
The Yuzu is worthy of trial as an acid fruit for
the home garden in subtropical or warm-temperate climates that are too cold to
permit the growth of other acid citrus fruits. The Yuzu was
formerly used widely in Japan as a rootstock for the satsuma orange, but now
the trifoliate orange is almost exclusively used in nurseries which propagate
the satsuma on a large scale, doubtless partly because seeds of the Yuzu are
only to be obtained in moderate quantity from fruits that have a market value,
whereas trifoliate orange seeds can be had in large quantity from fruits that
are of no value.
Nagai and Takahashi (1928,
pp. 2018-22, and 1925 [in Japanese]) found that of the five following
rootstocks—Yuzu, sweet orange, sour orange, trifoliate orange, and Japanese
summer orange (Natsudaidai)—the Yuzu gave the best results when approach-grafted
in order to rejuvenate Thomson navel trees on trifoliate orange
rootstocks. The sweet orange did nearly as well, but it is subject
to foot rot, a disease that does not attack the
Yuzu.
Shangyuan, or Ichang lemon [Citrus ichangensis
X C. grandis var. ?].
Illus.
Swingle, Jour. Agr. Res. 1:figs. 2-3. 1913; Hu, Calif. Citrog.
16:544, fig. 18. 1931; idem, Jour. Agr. Ass. China Nos.
126-127:61, fig. 29. 1934.
Distribution.—China: Lower valley of Yangtze River in Hupeh, Anhwei,
Kiangsu, and Chekiang provinces; sparingly cultivated.
An erect, much branched tree with lower branches
short, giving the tree a columnar outline; twigs glabrous, spiny; leaves ovate,
ovate-lanceolate, or ovate-acuminate, apices sometimes minutely emarginate,
sometimes bluntly pointed, margins obscurely crenulate, leaf blades about 3
times as long as the petioles, 6-9 X 2.5-3.5 cm, glabrous, tips rounded,
margins subentire; petioles more or less broadly winged, 20-25 X 6-12 mm;
fruits large, 8-11 X 7-10 cm, broadly obovate, usually with a low mammillate
tip surrounded by a shallow furrow; peel thick, 7-10 mm, lemon yellow when
ripe, very fragrant; oil glands concave; segments 10 or 11 with numerous large
seeds, 4-6 per segment, usually 16-20 (sometime longer) X 9-12 X 7-10 mm,
tapering into a flattened truncate base, sometimes polyembryonic; pulp very
acid, but with a strong, aromatic aftertaste; pulp-vesicles fusiform, 8-10 X
2-4 mm, on stalks 2-8 mm long, with a central streak of minute granules of acrid
oil.
The Shangyuan, or "Ichang lemon," which
was introduced into the United States about 1914, has proved to be very
resistant to winter cold. In southern Alabama it has endured
without harm temperatures that injured or killed most of the satsuma orange
trees of the same age growing near by, although the satsuma is a very
cold-resistant variety. At first the plant was supposed to be C.
ichangensis but was soon found to be very different, as it has very much
larger fruits and very much smaller winged petioles; it is, in fact, a
hybrid. There is some variation among different trees of the Ichang
lemon growing in China, such as is found among other violent hybrids of
Citrus.
Hu in 1930 named this hybrid
Citrus grandis var. Shanyuan; the next year he corrected the
spelling of the varietal name to read Shangyuan (1931, p.
544). He noted the similarity of this tree to the pummelo but stated
that the fruits are smaller and have a bitter taste. He considered
it to be a hybrid of the pummelo. He also reported (1930) that it
is grown in the lower valley of the Yangtze River, in Anhwei, Kiangsu, and
Chekiang provinces; later (1931) he stated that the fruit, though ornamental,
is not eaten and that the tree is hardy enough to be grown in Nanking.
Hu's varietal name for this hybrid is a phonetic
rendering of the Chinese name for it, namely, Hsiang yüan, or "Fragrant
ball," an allusion to its pleasing aroma. Tanaka (1930a, p.
32) changed Hu's name to a botanical one, Citrus hsiangyuan
Hort. Later he named the hybrid Citrus wilsonii
(1932b, p. 37), but stated: "The present species unquestionably has
Citrus junos in one parent and seems to be a cultigen originated through
the chance seedling."
There is no valid
reason for calling this plant a botanical species, as it is undoubtedly a chance
hybrid that arose under cultivation. Its resemblances to the Yuzu,
remarked by Tanaka, are doubtless due to the fact that both the Yuzu and the
Ichang lemon, or Shangyuan, are hybrids of C. ichangensis with two
different species of the subgenus Citrus. Fortunately, the
International Code of Botanical Nomenclature (Lanjouw, 1961, p. 31) now
gives a very simple method of citing such sports and cultigens under the name
of the species from which they arose by giving the variety name in some living
language (not in Latin) following the species name and surrounded by single
quotes. Under this rule any cultivated form of a species of
Citrus can be cited after the species name under its common name; for
example, C. sinensis 'Hamlin' or C. reticulata
'Dancy.' Many of the complex hybrids of Citrus can best
be handled in this way until their origin is worked out experimentally.
In China, the ripe fruits of the Ichang lemon
are used to perfume rooms or cabinets. In the United States, the
fruit has been occasionally used to make lemon pie, the heat of baking evidently
driving off the acrid oil that gives a disagreeably strong odor to "lemonade"
made from the juice; some who have made pies from Shangyuan juice prefer them
to regular lemon pies.
The Ichang lemon is a
very hardy plant and deserves trial as a rootstock in this country, where it
grows vigorously. It produces very numerous large seeds that
should make it easy to grow in the nursery. It also produces pollen
freely and hybridizes spontaneously with the satsuma orange if growing near by;
doubtless it could be crossbred with other forms of citrus fruits as
well. It should be used in breeding new hybrids for trial as
rootstocks or as acid fruits for home use in the cotton belt.
12. Citrus latipes (Swing.)
Tan. Studia Citrol. 2:155. 1928. Citrus ichangensis subsp.
latipes Swing. Jour. Agr. Res. 1:12. 1913. Illus. Tanaka,
Kankitsu No Kenkyû (Citrus Studies), 79, fig. 72, also 167, fig. 136(1). 1933;
fig.
3-48 this work.
Type.—Northeastern
British India, Khasi Hills, Living Bridge (Hooker f. and Thomson).
Herb. Kew.
Distribution.—Northeastern
India: Khasi Hills; northern Burma; grows in the mountains at considerable
elevation, 500-1,830 m [1,640-6,000 ft.].
Common name.—Khasi papeda.
A thorny tree
similar to C. ichangensis but having leaf blades more variable in size
and shape and with the tips subacute or even bluntly rounded, not apiculate or
subcaudate with blunt points as in C. ichangensis. The
flowers, instead of being borne singly in the axils of the leaves as in C.
ichangensis, are sometimes, at least, borne in small axillary racemes with
5-7 flowers. These latter flowers are much smaller, are 4-merous
(instead of 5-merous as is the large single flower of C. ichangensis),
and all the parts are smaller. However, the fruits are borne singly
and resemble those of C. ichangensis except for having a thicker peel, of
which the inner layer is chalky white just below the outer green
layer. The seeds are also smaller and more numerous than those of
C. ichangensis and are arranged 5-7 in each segment.
The following is a detailed description of
flowers and fruits (after Tanaka, 1928a, p. 155): Inflorescences
racemose, having 5-7 flowers; peduncles short, 5-8 mm, pubescent; pedicels 7-8
mm long, moderately thick, becoming thicker at the upper end where they join
the calyx, striated, glabrous; flower buds medium-sized, ovoid, about 7 X 9 mm,
rounded at the apex, showing oil glands indistinctly at the surface; calyx
discoid, 4 mm diam., lobes 4, broad, semicircular, wrinkled, slightly convex at
the tip and curving outward, rather thick, slightly pubescent at margins,
glabrous above; flowers, when open, 1.5 cm diam.; petals 4, spreading
horizontally, thick, oval; stamens 18-20, 7-8 mm long and about 1 mm wide,
filaments almost free at the tip but arranged in 6-7 groups each composed of
3-4 united together at the bases, thick, flat, wrinkled, pointed at the tip;
anthers oval or elliptic-oval, about 1.5 mm long, pointed, with small gland at
the apex; pistils shorter than stamens, about 7 mm long; ovaries
depressed-globular, 2 mm diam.; styles rather stout, 2.5 mm long, 1 mm diam.,
distinctly articulated with the ovary; stigmas large, depressed-globose,
1.5-2.5 mm diam., with 4 sometimes indistinct furrows; fruits medium-sized,
almost globular, slightly compressed at both ends; oil cells moderately small,
more or less equal in size, rather dense, convex; peel somewhat thick, composed
of rather dense tissue; central column (axis) of even width, moderately large;
segments 9, fairly large, outer margin very much rounded, inner margin also
rounded; pulp-vesicles few, fairly large, spindle-shaped, not short, relatively
well developed with a thick membrane, stalks very short; seeds numerous,
arranged 5-7 in each row, rounded, moderately large, parallel and
horizontal.
This species was founded on plants
collected in the Khasi Hills of northeastern India, but very similar forms are
found in the mountains of northern Burma. The type specimen in the
Kew Herbarium on which Swingle based C. latipes (1913c, p. 12) has
a fruit in cross section measuring 5 cm in diameter, peel 5 to 6 mm thick, with
nine segments.
In view of the extreme
cold-resistance of the typical C. ichangensis (and its hybrid, the
so-called Ichang lemon) it would be well worth while to introduce C. latipes
into the United States and see if it also is hardy and perhaps of value for
use in breeding new hardy acid citrus fruits and hardy rootstocks.
Subgenus Papeda Section Papeda
Subgenus Papeda Section Eupapeda
Swing. in Webber & Batchelor, Citrus Indus. 1:432. 1943.
Flowers small (less than 2 cm, often less than
1.5 cm diam.); stamens free from the beginning; winged petioles broad, and more
or less triangular; seeds small or medium-sized, not angular, much longer than
broad.
The species of the section
Papeda of the subgenus Papeda are remarkable in having very large
winged petioles and small flowers (often very small) with free staminal
filaments. The pulp-vesicles contain numerous droplets of acrid,
bitter oil that gives the pulp a very disagreeable taste. The
pulp-vesicles are often attached not only to the dorsal walls of the locules,
but also to the radial walls for one-half to three-fourths the distance from
the dorsal wall to the central core of the fruit. In this
character these species approach Clymenia.
As a result of the widespread use of the fruits
by natives, these species have come into culture on a small scale about villages
throughout the East Indies and in Melanesia and Polynesia. In this
way these species have been exposed to cross-pollination not only with other
species of the Papeda group but also with edible-fruited species of the
subgenus Citrus grown as dooryard trees.
Rumphius, near the close of the seventeenth
century, described and figured many forms of the Papeda group found
growing in Amboina and in other islands of the East Indian Archipelago; his
observations were published by Jan Burmann in 1741-1755 (Herbarium
Amboinense, vols. 2 and 5). Wester (1913; 1915, 1917a,
1917b, and 1924) published good descriptions and photographic
illustrations of many forms of the section Papeda group found growing
wild or cultivated on a small scale in the Philippine Islands, especially in
the southern islands, Bohol, Cebu, Negros, and Mindanao. Several
forms are likewise found in the southern half of Luzon, the northernmost
island.
It is clear from the works of Rumphius
and Wester that the species of the section Papeda have been very
extensively hybridized, almost as much so as those of the subgenus
Citrus.
However, many of the species of the
section Papeda group still occur in a truly wild state in the primeval
forests of southeastern Asia, the East Indies, New Guinea, and neighboring
islands. In these species and varieties it is thus possible to see
the unobscured results of many million years of evolution, whereas in the
edible-fruited species, varieties, and forms belonging to the subgenus
Citrus the results must be surmised rather than directly
observed. The papedas, because of their acrid, disagreeable flavor,
have not been considered by citrus experts of sufficient value to merit
study. However, hybrids between the species of the subgenus
Papeda and species of the subgenus Citrus have been known to yield
rootstocks very well adapted to support the commonly cultivated citrus fruit
trees. Several of the wild species of papedas are very vigorous and
deserve trial as rootstocks for the edible species of citrus.
Nothing is known about the vitamin C content of the papedas or about which
glucosides, if any, they contain.
13.
Citrus micrantha Wester, Phil. Agr. Rev. 8:20, pls. 5c,
6b. 1915. Citrus macroptera var. micrantha (Wester) Tan.
Trans. Nat. Hist. Soc. Formosa 22:430. 1932. Illus. Wester,
loc. cit. pls. 5c, 6b, and Phil. Bur. Agr. Bul.
27:pl. 12. 1913.
Type.—Philippines,
Bohol (Wester, No. 4829). Herb. Bur. Sci., Manila.
Distribution.—South Philippines.
Common name.—Small-flowered papeda.
Native name: Biasong.
The original description
of this species by Wester reads as follows: "A tree attaining a height of 7.5
to 9 meters, with comparatively small but sharp spines; leaves 9 to 12
centimeters long, 27 to 40 millimeters broad, broadly elliptical to ovate,
crenate, rather thin; base rounded or broadly acute; apex acutely blunt
pointed; petioles 35 to 60 millimeters long, broadly winged, up to 40
millimeters wide; wing area sometimes exceeding leaf area; flowers small, 12 to
13 millimeters in diameter, white, with a trace of purple on the outside, 2 to
5, in axillary or terminal cymes; petals 4; stamens free, equal, 15 to 17;
ovary obovoid, locules 6 to 8; style slender; distinct; fruit 5 to 7
centimeters long, 3 to 4 centimeters in transverse diameter, averaging 26 grams
in weight, obovate to oblong-obovate, somewhat compressed towards base; apex
blunt pointed; surface fairly smooth or with transverse corrugations, lemon
yellow; skin comparatively thick; pulp rather juicy, grayish, acid; aroma
similar to that of samuyao; juice cells short and blunt to long, slender
and pointed, sometimes containing a minute greenish nucleus; seeds many, flat,
pointed, more or less reticulate."
This
species stands out from all others of the subgenus Papeda because of its
very small 4-merous flowers, only 1.2-1.3 cm diam. when open. The
fruits are oval or obovate, blunt-pointed at both ends, 5-7 cm long but only
3-4 cm wide, with very few locules (only 6-8). The leaves have
broadly ovate blades, blunt-pointed at both ends, 3.5-6 cm long, 2.7-4 cm
wide. The broadly winged petioles, measuring 3.5-6 X 3.5-4 cm, have
a nearly straight or slightly curved top and gradually curved sides, tapering
to a blunt base. Seeds many, flat (2.5-3 mm thick).
This species is sparingly cultivated in the
southern Philippine Islands (Cebu, Bohol, Negros, and Mindanao), where it is
called biasong. Wester stated that the fruit of this species
is used as a hair wash but is not eaten and is of no economic importance.
13a. Citrus micrantha
var. microcarpa Wester, Phil. Agr. Rev. 8:21, pl. 7b.
1915. Illus. Wester, loc. cit. pl. 7b, and Phil. Bur. Agr.
Bul. 27:pl. 14. 1913.
Distribution.—Southern Philippines: Cebu and Bohol.
Common name.—Small-fruited
papeda. Native name: Samuyao.
The original description of this variety by
Wester reads as follows: "A shrubby tree, 4.5 meters tall, with slender
branches and small, weak spines; leaves 55 to 80 millimeters long, 20 to 25
millimeters broad, ovate to ovate-oblong or elliptical, crenulate, thin, of
distinct fragrance, base rounded to broadly acute; apex obtuse, sometimes
notched, petioles 20 to 30 millimeters long, broadly winged, about 14
millimeters wide, wing area somewhat less than one-half of the leaf blade;
flowers in compact axillary or terminal cymes, 2 to 7, small, 5 to 9
millimeters in diameter, white, with trace of purple on the outside; calyx
small, not cupped, petals 3 to 5; stamens 15 to 18, free, equal; ovary very
small, globose to obovate; locules 7 to 9, style distinct; stigma small, knob
like; fruit 15 to 20 millimeters in diameter, roundish in outline; base
sometimes nippled; apex an irregular, wrinkly cavity; surface corrugate,
greenish lemon yellow; oil cells usually sunken; skin very thin; pulp fairly
juicy, acid, bitter with distinct aroma; juice cells very minute, blunt,
containing a small greenish nucleus; seeds small, flattened, sometimes
beaked."
The samuyao was found by West
and Brown (1920, pp. 208-09) to yield from the rind "a clear, almost colorless
oil which is very fragrant. It should be useful as a perfumery
oil." The peel yielded 1.29 per cent of oil. They
stated that the crushed fruits are used by the women of Cebu for cleansing the
hair and are added to coconut oil to give it fragrance when applied to the
hair. The tree bears within five years from planting and produces
fruits during the entire year, but more abundantly during the rainy
season. West and Brown (1920, fig. 70) show the fruits as
distinctly pyriform, broadly rounded at the tip and tapering into a narrow,
conical base.
This variety has the smallest
fruit and the smallest flowers of any True Citrus Fruit Tree known but
curiously enough has more segments in the fruit (seven to nine) than C.
micrantha (six to eight), although the fruits of the latter are much larger,
5 to 7 by 3 to 4 cm, instead of 1.5 to 2 cm, as in the variety
microcarpa.
Wester stated (1915, p.
22) that in Bohol Island, P.I., "a somewhat more vigorous variety of the
samuyao was found that is called samuyao-sa-amoo" which has
"fruits a little larger, and smoother, and longer than broad, otherwise similar
to the samuyao." This description calls to mind Rumphius'
Limonellus aurarius, the goldsmith's lime, which occurs sparingly in
Amboina, Moluccas, and Ternate, a few hundred miles directly south of
Bohol. According to Rumphius (1741, vol. 2, p. 109, pl. 30), the
fruits of this plant are the smallest of all the limes (his figure shows ovoid
fruits 2.5 to 3 by 2 to 2.5 cm) and the leaves are also small, only 3 inches
long in all, about one-third of which represents the cordate, winged
petiole. The fruits are not eaten but are used by goldsmiths to
clean gold objects. The fruits also enter into native
medicines.
Both C. micrantha and its
variety microcarpa are of much scientific interest, as they are perhaps
the most primitive members of the subgenus Papeda, itself the most
primitive group of species of the genus Citrus.
14. Citrus celebica Koord. Fl.
N. O. Celebes, Mededeel. uit 's Lands Plantent. 19:639, also 370. 1898.
Illus. Koorders, Suppl. Fl. N. O. Celebes 2:pl. 19. July, 1922;
loc. cit. 3:pls. 10-11. December, 1922.
Type.—Northeastern Celebes
(Koorders). Hort. Bot. Buitenzorg, Java (now Kebun Raya, Bogar,
Java).
Distribution.—Northeastern
Celebes, southern Philippines.
Common
name.—Celebes papeda.
Leaves with small,
lanceolate, subcrenulate blades, 4.5-5.5 X 2.5-3.5 cm; winged petioles larger
than the blades, 5-6.5 cm long, 3-3.4 cm wide, oblanceolate, with a
subcordate or slightly curved upper end and gradually rounded sides tapering
into a deltoid base, having from 5-10 mm of the petiole almost wingless;
margins faintly crenulate; flowers occurring in 3- to many-flowered spikes or
small cymoid clusters in the axils of the leaves; flowers medium-sized, 15-17
mm when open, 4-5-merous; ovary reversed-pyriform, tapering sharply below,
broadly rounded at the tip with many (17-20) locules; style short, 5-6 mm;
stamens short, free; fruits large (10 cm diam.), globose, with a very thick
peel (about 3 cm); seeds small, probably monoembryonic.
A small tree 5 m high, growing in a wild state
in primeval forests at Karowa, in extreme northeastern Celebes (Minahasa) (Lat.
1° 15' N., Long. 124° 45' E.); fruits inedible.
The original description of this species
published in 1898 was not adequate to show its nature clearly, as the
description did not give the number of locules in the ovary, an important
character in the subgenus Papeda. Twenty-four years later
C. celebica was fully figured; then it became clear that this species was
distinct from C. macroptera.
14a. Citrus celebica var. southwickii (Wester)
Swing. Jour. Wash. Acad. Sci. 28:533. 1938. Citrus southwickii
Wester, Phil. Agr. Rev. 8:16-17, pls. 3c, 4c. 1915.
Illus. Wester, loc. cit. pls. 3c, 4c, and Phil. Bur.
Agr. Bul. 27:pl. 16b. 1913. (fide Wester, loc. cit.
1915 = C. southwickii).
Type.—Philippines, Bohol (Southwick, Wester). Herb. Bur.
Sci., Manila.
Distribution.—Philippines: Bohol and Mindanao.
This plant appears to be a variety of Citrus
celebica, with which it agrees in having very numerous locules (15-19) in
the ovary. It also agrees well in flower characters, especially in
the short style and the ovary tapered toward the base, but differs in having
smaller fruits with thin peel, and in having the blade of the leaves decidedly
larger than the winged petioles and often much blunter-pointed. The
winged petioles are similar to those of C. celebica but decidedly
smaller. Both the species and the variety have smooth
fruits. The difference in the diameters of the fruits is
misleading, since the fruits of variety southwickii, because of their
thin skin, have more pulp than the decidedly larger but very thick-skinned
fruits of the species.
The variety
southwickii was found by E. F. Southwick in Bohol, where it is not
uncommon and where it is cultivated. It was also found by Wester in
Baganga (Lat. 7° 35' N., Long. 126° 35' E.), in eastern Mindanao, about 450
miles north-northeast of Karoa in northern Celebes. It has been
introduced into the United States.
It is
possible that this plant may prove, upon more detailed study, to be a complex
hybrid of C. celebica with one or two other species. The
thin, smooth skin and the abundant, though acrid, pulp and the short,
broad-winged petioles arouse suspicion that it may be a complex hybrid of C.
celebica with a pummelo (C. grandis), or with some other species of
the subgenus Citrus.
This variety is
promising for breeding purposes because of its fair-sized, juicy, smooth,
thin-skinned fruits. The bitter taste of the pulp, due to the acrid
oil in the pulp-vesicles, can doubtless be removed by hybridization with the
species of the subgenus Citrus that have little or even no oil in the
pulp-vesicles, such as C. reticulata. It is highly probable,
even if variety southwickii proves to be a dilute hybrid of the pummelo,
that wild forms of C. celebica will be found native in the southern
islands of the Philippines, since southern Mindanao is only about 300 miles
north of northeastern Celebes; Bohol, where variety southwickii occurs,
is less than 400 miles distant.
OTHER POSSIBLE HYBRIDS OF CITRUS CELEBICA
Alemow.
This hybrid, named C. macrophylla by Wester (1915, p. 16, pls. 3b,
6c), is sometimes cultivated in Cebu, P.I. It has large
leaves, with the blades 12 to 14 cm long and 6 to 8 cm wide, with much smaller,
subtriangular, short-winged petioles, measuring up to 3.5 cm wide near the
top. The fruits are very large, 8.5 to 10 cm in diameter,
subglobose to oblong, more or less narrowed at the base, with a rough,
transversely-corrugated, but rather thin skin. The fruit has 13 to
16 segments and rather dry, sour pulp, considered inedible even by the
natives.
This seems to be a hybrid of C.
celebica, or some other species of the subgenus Papeda, with a
species of the subgenus Citrus, probably a pummelo (C.
grandis). It has shown some promise as a rootstock for lemons in
California.
Kabuyao.
This is another possible hybrid of C.
celebica. It is called kabuyao in Luzon Island, P.I., and
kopahan in Bohol Island. It was described by Wester (1915,
pp. 17-19, pl. 5b, fig. 1; and 1924, p. 93, pl. 31c) as typical
C. hystrix. It differs from that species, however, in having
more numerous stamens (30 to 36) and more locules (13 to 18) in the
ovary. The fruit is subglobose or short-pyriform, 7 to 9 cm long
and 7 cm in diameter, with a smooth skin; the pulp-vesicles contain a
"nucleus," doubtless composed of droplets of acrid oil such as are found in all
species of the Papeda group and in most of their hybrids.
Amongpong.
This plant is found in Bohol Island, P.I., and is
evidently similar to the kabuyao, but according to Wester (1915, p. 18)
it has only 26 to 30 stamens and a larger fruit, 10 cm in diameter.
These two hybrids, the kabuyao and the
amongpong, and other similar fruits may perhaps be hybrids of C.
celebica (which has many locules in the ovary) with C. macroptera or
with C. hystrix, both members of the subgenus Papeda.
The increased number of stamens and the larger fruits might result from such
hybridization.
15. Citrus macroptera
Montr. Mém. Acad. Sci. Lyon 10:187. 1860. Citrus papuana F. M.
Bail. Contr. Fl. Brit. N. Guin. 1903; C. aurantium subsp. saponacea
Safford, Contr. U.S. Natl. Herb. 9:226. 1905. Illus. Bailey,
loc. cit. (text fig.); Tanaka, Kankitsu No Kenkyû (Citrus Studies), 131,
fig. 106. 1933 (?); fig. 3-49 this work.
Type.—New Caledonia, Art Island (Montrouzier). Herb. Fac.
Med. Pharm., Univ. Lyon, France.
Distribution.—Thailand, Indo-China, Philippines, New Guinea, New
Caledonia, and Polynesia (possibly escaped from culture here).
Common name.—Melanesian papeda.
Father Montrouzier's original description in
Latin may be translated as follows: "Petioles broadly winged, distinctly
articulated; leaves elongate-acuminate, only twice as long as the petiole,
subcrenulate, with very numerous [oil] glandlets, 5-6 inches [12-14 cm] long
excluding the petiole; twigs subcompressed, with one long spine on young twigs
and one short spine on old twigs in the axils of the leaves; peduncles
axillary, often extra-axillary, several crowded together, bracteate; calyx with
4-5 small sepals; corolla with 4-5 petals with valvate aestivation; petals
elongate, concave; stamens 20; ovary subhemispherical; style thick, round;
stigma depressed; fruit with 10-12 segments, pubescent; segments 1-2 seeded,
with scanty pulp, depressed, almost without juice, about the size and form of
an orange. A tree 15-16 feet [4.2-5 m] high, growing near the
houses of the natives on the Island of Art. It flowers in
September, and is commonly called don gan."
Bailey's original description of his Citrus
papuana, doubtless a synonym of C. macroptera, reads as follows:
"Branchlets green, very angular like those of the common orange, all parts more
or less covered with amber-coloured lenticular glands; axillary spines
straight, erectopatent, about 1/2 in. long. Leaves
broadly-lanceolate, 2 to 5 in. long, the winged petiole cuneate 2 to 3 1/2 in.
long and from 1 to 2 in. broad at the top; margins with broad shallow
crenulations, the lenticular glands very numerous on the midrib, both surfaces
punctate; lamina very freely disarticulating from the petiole. (No
flowers collected.) Fruit globose, about 2 1/2 in. in diameter,
pale yellow, smooth; glands small, slightly concave; ring about 1/2 in. thick;
cells 10. In foliage this species somewhat resembles C.
hystrix, D.C."
This species was discovered
by Father Montrouzier on the Island of Art, situated a few miles to the
northwest of the north end of New Caledonia. What appears to be the
same species was described and figured in 1903 by F. M. Bailey from Milne Bay,
New Guinea, about 1,200 miles to the northwest. Unfortunately
Bailey had no flowers and Montrouzier in his account does not describe the
flowers in adequate detail.
This species is
characterized by large leaves, sometimes 10 or 12 inches long (including the
very large winged petiole) and 2 inches wide; it has subglobose fruits the size
and shape of an orange, but with very little juice.
Wichmann (1917, p. 202) told of getting a fresh
supply of fruits at a village on the shores of Lake Sentani in northern Dutch
New Guinea (now West Irian, Indonesia). Among these were
thick-skinned fruits of a Citrus with pulp "so disgustingly bitter that
it was hours before one's mouth got free from the taste." This
persistent bitter taste is due to the numerous droplets of acrid oil that are
found in the pulp-vesicles of all species of the subgenus Papeda and in
some of the hybrids of them with species of the subgenus Citrus.
A beautiful Citrus which Swingle saw
growing wild in the primeval forest on the lower slopes of Mount Maquiling in
southern Luzon, P.I., belongs, he believed, to C. macroptera.
It has usually long-pointed leaves with very large, entire, winged
petioles, and smooth, subglobose fruits the size of a large
orange. The conviction that this Citrus is truly indigenous
in the primeval forests on the lowers slopes of Mount Maquiling is
substantiated by the observation of D. A. Herbert (1924, p. 195), who reported
that this Citrus (which he called C. hystrix) occurs occasionally
in the dipterocarp forest, which was the original forest growth on Mount
Maquiling up to an altitude of about 600 meters.
This species doubtless grows wild in New Guinea,
Bismarck Archipelago, Celebes, southern Luzon, and the southern islands of the
Philippines. It has reached New Caledonia, Fiji, Samoa, Guam, and
probably many other of the Polynesian islands, probably being carried by the
Polynesians for use in washing. Safford (l.c., pp. 226-27) gave a
firsthand account of the use of the fruit pulp of this species in Guam not only
for washing the hair, but also as a substitute for soap in washing
clothing. He stated further that the macerated leaves form a lather
when water is added. Safford whose observations apply also to
Samoa, cited F. Reinecke (1898, pp. 642-43) to show that both the pressed-out
fruit juice and the macerated leaves form a lather when rubbed and that they
are specially useful in washing the hair to free it from the lime which is much
used by the natives to bleach the hair. Both Safford and Reinecke
noted that after the fruit falls off the tree the peel hardens and becomes like
a stony shell in drying.
Of all the species
of the Papeda group, C. macroptera is probably the most promising
for use as a rootstock. F. M. Bailey (1903, p. 1) was so impressed
with the vigor of the specimens of this species from Milne Bay, New Guinea (his
C. papuana), that he recommended it as a rootstock for cultivated citrus
fruit trees in tropical Queensland. Hybrids of C. macroptera
with vigorous species of the subgenus Citrus might be expected to
show still greater energy of growth.
The
plants of C. macroptera that have been grown in Florida from seed
obtained from trees found growing wild on Mount Maquiling, Luzon, P.I., were
reported by T. Ralph Robinson (Keys, 1923) as being immune to withertip and
resistant to canker. This large-fruited, disease-resistant, and
vigorous strain is promising for use in breeding new and superior citrus
rootstocks.
HYBRIDS OF CITRUS MACROPTERA
Moli
kurikuri hybrid.
A hybrid of C.
macroptera, very probably with the pummelo (C. grandis), was named
C. vitiensis by Tanaka (1928b, p. 715). It was found
in Fiji in 1860 by Berthold Seemann (1862), who collected it together with the
parent species, C. macroptera, giving both specimens the same number
(58). Both sheets are preserved in the Kew Herbarium.
One of the specimens is the typical C. macroptera Montr., the moli
kurikuri of the natives, said by Seemann to be a wild orange, found on the
island of Viti Levu, that is not eaten but is "used to kill lice by washing the
hair." The manuscript note on the moli kurikuri was
attached, doubtless by accident, to the sheet on which was mounted the
specimen of the hybrid, which shows the following characters: twigs terete,
internodes short (1.5 to 3.5 cm); spines straight, acute, short (only 3 to 4 mm
on mature branchlets); leaf blades narrow, lanceolate, acuminate or caudate at
apex, broadly rounded at base, 4.5 to 5 by 1.8 to 2.5 cm, margins shallowly
serrate, veins numerous but obscure; petioles winged, obpyramidal, with
slightly curved tips curving abruptly and nearly straight-sided, cuneate bases
20 to 26 by 10 to 15 mm. Seemann's manuscript note reads further:
"it was plentiful on the banks of the Namosi river growing in company of
shedocks28 and flowering together
with…[that] species." This circumstance would easily account for
the finding of a hybrid between these two species! This hybrid has,
in fact, leaves somewhat like those of the so-called Shangyuan, or Ichang lemon, which is doubtless a hybrid of C. ichangensis
(also of the subgenus Papeda) and C. grandis.
To name as a valid new species a chance hybrid
found growing in immediate proximity to both of its parents is to give unmerited
importance to such a plant.
Webber's
Philippine hybrid.
Webber's Philippine
hybrid is a mandarin-like citrus fruit tree which was named C. webberii
by Wester (1913, pl. 14; 1915, p. 13; 1917, p. 108; 1924, p. 96, pls.
20b, 30a) in honor of his former chief, H. J. Webber.
It is called kalpi in the Bicolano dialect. According to
Wester (1924, p. 96), it is a medium-sized, handsome tree 5 to 10 meters tall,
bearing very juicy, acid fruits that can be used like lemons. It
grows in both the large islands of the Philippines, Luzon and
Mindanao. The fruit is variable in size, depressed-globose, with 9
to 11 segments; skin very thin, yellowish when ripe. Wester (1917,
p. 107, pls. 1, 3, 4) reported that the kalpi when used as a rootstock
appears more congenial and produces a more rapid growth than other of the
Citrus species of Philippines origin. It seems to be a hybrid
of the native C. macroptera with the common Philippine mandarin (see the
Chinese Yuzu,
a similar hybrid of the papeda with a mandarin).
Another form of Webber's Philippine hybrid is a
tree with weeping branches considered by Wester (1917, p. 108, pl. 1a)
to be "the most ornamental species in the genus Citrus." It
grows well at the Citrus Research Center at Riverside, California, where its
handsome shape and beautiful foliage make it a perpetual tribute to Dr.
Webber. It should also be tested as a rootstock, as it produces
seeds abundantly.
Kansi.
A Philippine citrus called kansi by the
natives of Bohol, where it is sparingly cultivated, may be a hybrid of C.
macroptera, which occurs in Bohol, with a pummelo (C.
grandis). The kansi has leaf blades broadly ovate to
elliptical, pointed at the tip, broadly acute at base, with subentire margins,
9 to 12 by 3 to 4.5 cm; winged petioles broadly obovate, even-margined, 3.5 to
4.5 by 2.5 to 3 cm, having less than half the area of the leaf blade; stamens
20 to 23, free; ovary oblate; fruit oblate, smooth, lemon yellow, 38 to 40 by
44 to 46 mm, pulp acid, peel 3 to 5 mm thick, segments 11 to 14, seeds many,
flattened, monoembryonic. This was called C. hystrix var.
boholensis by Wester (1915, p. 19, pls. 4, 5), who stated that the fruit
is eaten with fish and also makes a fairly good ade. That it is a
hybrid of C. macroptera with a pummelo is borne out by the statement of
Wester (1915, p. 12) that pummelos with from 11 to 14 segments in the fruits
are widely distributed in the Philippines.
15a. Citrus
macroptera var. kerrii Swing. Jour. Wash. Acad. Sci. 32:24.
1942. Illus. Swingle, loc. cit. figs. 1 and 2.
Type.—Thailand, Nakawn Sawan, Kampêngpat,
Mê Lamung (Lat. 16° 15' N., Long. 98° 58' E.), altitude about 500 m (Kerr, No.
6081, coll. June 7, 1922). Herb. Univ. Aberdeen.
Distribution.—Northern and central
Thailand; North Vietnam: Tonkin; southern China: southern Yunnan Province.
Common name.—Kerr's Thailand papeda.
Differs from the species in having larger fruits,
8-9 cm diam. instead of up to 5-6 cm, with more segments, 12-13 instead of 10
(rarely 12), with smaller, thinner-walled, ovoid pulp-vesicles attached in
large numbers to the side (radial) walls of the locules for 2/3-3/4 the
distance from the dorsal walls of the segments to the center of the fruit, and
with a thicker peel, 12-14 mm thick instead of 5-6 mm. The spines
on vigorous, young, vertically-growing shoots are very long (6-7 cm), straight,
sharp, arising nearly at right angles to the direction of the twig, but on
fruiting branches they are usually only 5-10 mm long or wanting
altogether. The leaf blades are sometimes ovate, rounded at both
ends, often emarginate at the apex, about 4-7 X 3-3.5 cm, with merely undulate
margins; petioles broadly winged, obovate, with slightly undulate margins, 5-7 X
3-3.8 cm, often slightly larger than the leaf blade. In another
collection from Ban Kragê, Thailand (Kerr, No. 11983), the leaf blades are 8-10
X 4-5 cm, broadly ovate-acuminate, tapering into a narrow acumen, blunt at the
very tip; petioles 5.5-7.5 X 3.5-4.5 cm, obovate, with crenulate or subserrate
margins. In material from Tung Kung in northern Tonkin (Groff, No.
19877), the leaf blades are lanceolate-acuminate, 6-8 X 2-3.5 cm,
crenate-denticulate above the middle, and tapering into a long, slender acumen
which often has a very sharp apex; petioles 5-7.5 X 2.2-4.4 cm, with entire or
slightly undulate margins. Flowers are lacking on all the material
examined except on Kerr's No. 11983, where they arise singly (or in 2's or 3's)
in the axils of the leaves on short, slender, glabrous pedicels, 3-5 mm long;
flower buds subglobose, 3-4 X 2.5-3 mm; calyx very small, saucer-shaped, lobes
4 (?), bluntly triangular, 1 mm long, 2 mm wide; petals 3.5-4.5 X 3-4 mm;
stamens numerous (16-20 ?) with slender free filaments. Several
specimens show fruits. They are subglobose, 8-9 cm diam., with
thick peel (1-2 cm) almost entirely composed of white spongy tissue with a
superficial green layer, only about 1 mm thick, which contains very numerous,
nearly spherical oil glands. The fruits show 12 and 13 segments
with lateral (radial) walls 2-2.5 mm thick where they join the peel, tapering to
0.8-1 mm where they join the clearly delimited cylindrical core.
The pulp vesicles are very numerous, obovoid, very small, 1.5-2 mm long,
bluntly pointed, tapering into slender stalks, 1-3 mm long. The
pulp-vesicles are attached not only to the adaxial wall of the carpel but also
to the lateral (radial) walls 2/3-3/4 the distance from the dorsal wall to the
core. The seeds are immature in Kerr's specimens, but seem to be
placed almost exactly in the inner angle of the segments and to lie nearly
parallel to the core.
The type specimens of
this variety were collected in Nakawn Sawan Province of Thailand by A. F. G.
Kerr, who stated that it is a "common tree in evergreen forest, growing up to
10 m high, fruit 8 cm diam.; lower shoots armed with long spines."
Another of Kerr's specimens of this same number in the Aberdeen University
herbarium that Swingle had opportunity to examine through the courtesy of J. R.
Matthews shows a piece of a very angular, upright-growing new shoot having five
internodes, with five very strong, sharp, straight spines, 6 to 7 cm long, 4 to
5 mm in vertical diameter at the base and 2 to 2.5 mm in transverse
diameter. These spines arise almost exactly at right angles to the
twig. The winged petioles are very nearly the same length as the
spines that arise in their axils. They are 3 to 4 cm wide at the
very broadly rounded tips and taper to a cuneate base.
This variety shows some resemblance to C.
latipes, but apparently has decidedly smaller flowers and more globose
fruits with much thicker peel. More study is needed to allocate
this interesting papeda with certitude to its proper taxonomic position.
What seems to be a form of this variety with
acuminate leaf blades was found by the late G. W. Groff when at Lingnan
University, Canton, China, while making botanical exploration in cooperation
with the former Bureau of Plant Industry, U.S. Department of Agriculture, in
northern French Indo-China in January, 1932. Groff collected good
herbarium material (Lingnan Univ., No. 19877) from four trees (very like those
found by Kerr) growing at Tung Kung in Tonkin (about Lat. 22° 15' N., Long.
102° 50' E.) 130 kilometers (80 miles) west of Lau Kai (11-12 km northwest of
Cha Pa at Ta Ching on the forest road from Ku Ho to Ta Ching) on the
Yunnan-Tonkin border. The trees were growing in the heart of the
forest at an altitude of about 5,000 feet in rich moist soil.
15b. Citrus macroptera
var. annamensis Tan. Bul. Mus. Hist. Nat. Paris, 2 sér. 2:164.
1930.
Type.—French Indo-China, Co-inh
Mountains, Nhatrang, Annam (Poilane, No. 4650). Herb. Mus. Hist.
Nat., Paris.
Distribution.—Known only
from the type locality.
Common
name.—Annam papeda.
The original
description reads in translation as follows: "Leaves very large, leaf blades
ovate, acute at apex, obtuse at the base; petioles broadly winged; petiole
wings oblong, truncate, a little longer than the leaf blade. Fruits
medium-sized, ovate-obovoid, peel glabrous, thin; pulp-vesicles scanty, very
short."
A photograph of the type specimen of
this variety sent to Swingle by Tanaka shows the leaf blade to be broadly oval
or lozenge-shaped, 6-7 X 3.5-4 cm, bluntly pointed or subacute at the apex and
broadly cuneate at the base, with the upper half or two-thirds with undulate,
crenulate-dentate margins; petioles broadly winged, linear-oblong, decidedly
longer than the leaf blade but not any wider, 7-11 X 3.5-4 cm, truncate at the
apex and abruptly rounded at the corners into a more or less oblong section with
nearly parallel margins including 1/2-3/4 the distance to the base; the lower
1/2 or 1/4 of the petiole is gradually narrowed into a cuneate base, with a very
short segment of the petiole (3-6 mm long) at the base wingless; the fruit is
depressed globose, 4.5 cm high and 5.2 cm wide; peel 3-4 mm thick.
This variety is strikingly different from typical
C. macroptera in the oblong shape of its very long winged petioles, which
resemble those of some forms of C. ichangensis; the fruits, however, are
entirely different from those of this latter species, but are much like those of
C. latipes. This variety differs from C. macroptera
var. kerrii in having oblate fruit with rather thin peel instead of
globose fruits with remarkably thick peel, often five times as thick as that in
this variety.
According to Tanaka (1937, p.
238), this variety occurs also in Thailand, Burma, and northeastern India,
where the fruits are stored and "used for refreshing drinks when other small
acid Citrus fruits are gone." He did not hesitate to identify it
with the Citrus combara of Rafinesque (1838, p. 142), but the description
there given is inadequate and might apply to many forms of bitter orange
belonging to the subgenus Papeda. This and other forms of
C. macroptera should be studied carefully, as some of them may prove to
be distinct species. All of them should be tested as rootstocks for
the cultivated species and varieties of True Citrus Fruit Trees belonging to
the subgenus Citrus.
16.
Citrus hystrix ["Citrus histrix"] DC. Cat. Pl. Hort. Bot. Montp.
97. 1813. Citrus hystrix DC. Prodr. 1:539. 1824; Papeda
rumphii Hassk. 1842; Citrus papeda Miq. 1855; C. torosa
Blanco, 1873. Illus. Rumphius, Herb. Amboin. 2:pls. 26 (fig.
1), 27, 28. 1741; Risso & Poiteau, Hist. Nat. Orang. pl. 59 (col.).
1818-1822; Vidal & Soler, in Blanco, Fl. Filip. ed. 3. 4:38, pl. 408
("var. colobot"). 1880; Vidal, Sinop. Fam. Gen. Pl. Leñosas Filip., Atl. pl. 25,
fig. F. 1883; Bonavia, Cult. Orang. Lem. India Cey. Atl. pl. 225, figs.
A-K. 1890; Sawyer, Agr. Jour. India 5:pls. 36 (col.), 37, 38.
1910; Wester, Phil. Bur. Agr. Bul. 27:1, pl. 11. 1913; Ochse, Indische
Vruchten, fig. 107. 1927; idem, Fruits Fruitcult. Dutch East Ind. pl. 46
(col.). 1931; Redgrove, Gard. Chron. 3 ser. 89:149, fig. 76. 1931; fig.
3-50 this work.
Type.—Montpellier
(De Candolle). Herb. Delessert, Geneva, Switzerland (fide
Tanaka, 1931b, p. 11).
Distribution.—East Indian Archipelago, Ceylon, Burma, Malay Peninsula,
Philippines; widely naturalized, native habitat uncertain.
Common name.—Mauritius papeda.
J. J. Ochse (1931, pp. 117-18) gave the following
description of C. hystrix as it occurs in the Dutch East Indies (now
Indonesia): "Low tree or shrub, 2-12 m high; trunk crooked, asymmetric or
angular, rather thin, branched near the base; crown irregular, densely
branched; branchlets thin, when young compressed-acutangular, when older
terete, dark green, glabrous with scattered glandular dots, armed with axillary
spines; spines short, stiff, subulate, green with hard brown or orange-coloured
tips, obliquely erect, solitary, glabrous, 0.2-1 cm long.
Leaves alternate or biseriate, stalked unifoliate, orbicular-ovate or
ovate-oblong, lanceolate; base cuneate, obtuse or rounded, rarely subcordate,
apex obtuse, rounded or obtusely acuminate, often notched; patently
serrate-crenate, coriaceous, glabrous on both surfaces, above dark green,
shining, beneath light green or yellowish-green, dull, densely pellucid dotted,
fragrant when bruised, 3-15 cm long, 2.5-6 cm wide. Petiole from
0.3-0.5 cm above the base upwards with large, foliaceous wings; the winged part
obovate or obcordate-oblong, with an acute, cuneote, obtuse or rounded base and
an obtuse, truncate, rounded or slightly emarginate apex, patently
crenate-exsculptate, coriaceous, above dark green, shining, beneath light green
or yellowish green, dull, with scattered pellucid dots, including the wings 1-8
cm long, 1-4.5 cm wide, sometimes very similar to the leaf itself and not
rarely for a greater or smaller part connate with it.
Inflorescences axillary or terminal, dense, 1-5-flowered,
glabrous. Flowers smallish, shortly stalked, 4-5-merous,
fragrant, in bud globose, white, 0.5-0.7 cm diam.; pedicel terete, yellowish
green, glabrous, 0.1-0.5 cm long. Calyx cupular, afterwards
flat, 4-lobed, glabrous within and without, 0.2-0.35 cm diam.; lobes broadly
ovate-triangular, acute or shortly acuminate, yellowish white with violet
margins, ciliate, ±0.1 cm long. Petals 4-5, ovate oblong,
with a narrowed base and a narrowed or acuminate, obtuse apex, yellowish white
or tinged with red, glabrous, with scattered pellucid dots, 0.7-1 cm long,
0.3-0.5 cm broad. Stamens 24-30, quite free, 0.4-0.8 cm long,
filaments thickened above the base, with a filiform apex, glabrous, 0.3-0.6 cm
long; anthers elliptic-oblong, 0.15-0.25 cm long. Ovary
depressed globose, glabrous, with a tuberculate-folded surface, 0.2-0.4 cm
diam.; style short, robust, terete, glabrous, 0.1-0.15 cm long;
stigma depressed globose, glabrous. Fruit pendulous,
globose, ovoid or ellipsoid, with an abruptly contracted obtuse or rounded base
and a rounded or slightly depressed apex, crowned by a short style-rest, very
irregularly bumpy, glabrous, with many scattered glandular dots, when ripe
yellow or yellowish green, feebly shining, 5-7 cm diam.; peel thick, its
exterior layer of ±0.2 cm thickness, yellowish green, the inner part white;
pulp yellowish green, very sour and slightly bitter, faintly fragrant; fruiting
pedicel very short, 0.3-0.5 cm long. Seeds ovoid-oblong,
1-1.5 cm long, 0.5 cm thick; cotyledons and plumule white."
Ochse stated that "this species, called
Djerook pooroot in the Dutch East Indies, is nowhere cultivated on a
large scale but…very often on a small one."
This much misunderstood species was named by De Candolle from young seedlings,
not yet in flower, grown from seed sent to the Montpellier Botanic Garden from
Mauritius, where it was probably not native. De Candolle
redescribed it in 1824 but had not yet seen flowers or fruits.
Thanks to flowering material from Mauritius and
from Montpellier preserved in the Kew Herbarium, it is possible to determine
with certainty that C. hystrix is the species with very bumpy fruits
belonging to the subgenus Papeda that is widely distributed in
Indonesia. The fruits usually have 10 to 12 segments (rarely 13 or
14). The leaves are blunt-pointed, usually of medium size, 8 to 12
by 3 to 5 cm, more or less irregular at the tip and sometimes slightly
emarginate. The margins are more or less crenate. The
winged petiole, usually two-thirds to three-fourths as long as the leaf blade,
is broadly rounded and blunt at the base, often subtruncate at the tip, often
with more or less crenate margins.
The flowers
are small, with short, entirely free stamens, as in all the other species in
the section Papeda. The fruits are almost always bumpy or
tuberculate. The pulp-vesicles were first described by Penzig
(1887, pp. 131-32), who stated: "They are provided with a slender very long
stalk which suddenly enlarges toward the free end, forming a small round or oval
body pointed at the tip." Penzig noted that they differ from the
pulp-vesicles of the other species studied by him in showing "a large amount
of oil accumulated in their centers." This accumulation of numerous
droplets of oil, often of a very strong and acrid flavor, is characteristic of
all the species of the subgenus Papeda. The cotyledons are
epigeous in germination.
Citrus
hystrix, although the best known and most widely distributed species of
this subgenus, is not the most vigorous species, and not the most promising for
use as a rootstock. It has undoubtedly hybridized with other
species of the subgenus Papeda in the village fruit gardens of the
people of the East Indian Archipelago and also with species of the subgenus
Citrus. Doubtless some of these hybrids are included among
the bizarre forms described and figured by Wester (1913 and 1915).
Bonavia reported (1886, p. 215) that the fruit of
C. hystrix is never eaten; however, it is used by the natives in Ceylon
as an insecticide for washing the head and also for smearing on the feet to
kill land leeches. The tree is often cultivated near the villages
for these purposes. According to Bonavia, C. hystrix may have
come into Ceylon with the Dutch and their Malay servants for use as a hair
wash. This use may account for the wide distribution of the species
throughout the East Indian Archipelago and adjoining parts of the Asiatic
mainland.
HYBRIDS OF CITRUS HYSTRIX
Lemon
Martin.
The Limo tuberosus or Lemon
Martin of Rumphius (1741, vol. 2, pp. 101-03, pl. 26, fig. 1) is probably a
hybrid of C. hystrix with the lemon (C. limon). The
leaves are crenate-margined with broad, subcordate winged petioles, also
crenate-margined. The fruit is somewhat lemon-shaped but
exceedingly rough, with bosses or warts 5 to 10 mm in diameter. The
fruit figured by Rumphius is 5.54 by 4.5 cm and is said to be a yellowish-green
when ripe "like a Spanish lemon"; moreover the odor is somewhat like that of
the lemon. The juice is vinous, not very sour; according to
Rumphius, the fruits can be eaten raw, especially if sliced and sprinkled with
sugar. Rumphius stated that the Lemon Martin in his day (in the
last quarter of the seventeenth century) was believed to have been brought to
the East Indies by the Portuguese and to have been grown by Europeans in many
parts of the archipelago.
It seems very
probable that the Lemon Martin of Rumphius arose from lemons brought by the
Portuguese from Europe that were propagated from seed in the East Indies and,
through chance cross-pollination by insects, became hybridized with C.
hystrix, or with some other species of the subgenus Papeda.
Subtribe 3.
Balsamocitrinae: Hard-Shelled Citroid Fruit Trees
The subtribe Balsamocitrinae, the Hard-Shelled
Citroid Fruit Trees, is a remarkable side shoot of the orange subfamily not
very closely related to any of the other subtribes. It includes
seven genera occupying two noncontiguous areas of distribution.
Four genera occur in southeastern Asia from India to Burma and Indo-China,
reaching some of the neighboring East Indian islands, and also reaching Luzon,
the northernmost island of the Philippines. The other three genera
occur in tropical Africa, from Uganda west to the Ivory Coast and south to the
Republic of the Congo. None of the Hard-Shelled Citroid Fruits are
native to New Guinea, Australia, or the Polynesian islands in spite of having
spread from southeastern Asia as far as West Africa. This subtribe
is a side branch of the ancestral tree of the orange subfamily, a side branch
that parallels the subtribe Citrinae, the Citrus Fruit Trees. These
two subtribes resemble each other in having large fruits with hypermerous
ovaries (with 6 to 20 locules) with numerous ovules (6 to 16+) in each
locule. The genera comprising the subtribe Balsamocitrinae have no
pulp-vesicles, but they show a high specialization in the structure of the
fruits, which usually have a hard, almost woody exocarp (leathery in the genus
Swinglea). The locules are filled with resinous gum and the
seeds are sometimes sparingly hairy, woolly, or covered with laciniate
paleae.
The seven genera of this subtribe fall
into three groups. The first, group A (the Tabog Group), comprises
only one genus, Swinglea, native to Luzon, the northernmost island of
the Philippines. This genus has fruits so dissimilar in structure
to those of the genera in the other two groups that question may be raised
concerning its degree of relationship to them.
Swinglea, unlike the other members of the
subtribe Balsamocitrinae, has oval fruits with a leathery (not woody) shell
which has 8 to 10 longitudinal ribs (somewhat like a muskmelon) corresponding
with the number of locules. The outer wall of the fruit, which does
not separate like the peel of an orange, contains crowded, radially arranged
ellipsoid oil glands, 4 to 7 mm long. The locules contain many hairy
seeds immersed in a glutinous fluid apparently secreted by ellipsoid mucilage
glands, 5 to 10 mm long, which slant toward the axis of the fruit and also
toward the radial axis of the locule into which they open, as shown in figure 3-51.
The second set of
related genera (four in all) constitutes group B (the Bael-Fruit
Group). The best-known genus in this group is Aegle, native
to India, Ceylon, Burma, and Indo-China, and widely cultivated in the East
Indian Archipelago, especially Java. The other genera of this
group, Balsamocitrus, Afraegle, and Aeglopsis, occur
thousands of miles to the west, in tropical Africa, from Uganda west to the
Ivory Coast and southward to the Republic of the Congo and Angola.
It seems remarkable that those genera in group B which have the largest fruits,
obviously of common ancestry, were able to jump over the great distance from
southeastern Asia to West Africa, but failed to reach the Philippine Islands,
to say nothing of New Guinea and Australia.
Aegle and Balsamocitrus (both with trifoliolate leaves),
Afraegle (with normally trifoliolate leaves, reduced to unifoliolate or
to simple leaves on the older branches of A. gabonensis), and
Aeglopsis (with unifoliolate leaves) are obviously all closely related
but differ in important taxonomic characters, such as the number of locules in
the ovary and the number of stamens. Aegle has 8 to 20
locules in the ovary, Afraegle 6 to 8, Balsamocitrus 8, and
Aeglopsis 6 to 8 (sometimes 5). Aegle has woolly
seeds, whereas the other three genera all have glabrous seeds. The
development of the disk also varies greatly in the four genera: Aegle
and Balsamocitrus have a cylindric disk below the ovary and of about the
same diameter, whereas Afraegle and Aeglopsis have a greatly
enlarged and lobed disk that partly encloses the base of the ovary.
The third set of related genera, group C (the
Wood-Apple Group), comprises two genera, Feronia and Feroniella,
which occur in India, Burma, Indo-China, and Java. Feronia
limonia is often cultivated for its fruit. This group differs in
very important taxonomic characters from the other two groups, and is
undoubtedly one of great antiquity that now represents a side line of
development in the subtribe Balsamocitrinae.
Feronia and Feroniella, spiny trees with pinnate leaves and small
leaflets, have ovaries in which the locules (four to six) coalesce at an early
stage of development into a single cavity with numerous ovules borne on the
walls (parietal placentae), a character unique in the whole plant family
Rutaceae.
The evolution of the subtribes
Balsamocitrinae and Citrinae has doubtless proceeded along parallel lines from
a common ancestral form, both subtribes having attained nearly the same degree
of specialization. It is not possible, however, to exhibit in a
linear arrangement of the tribes, subtribes, and groups of the Rutaceae the
true status of the evolutionary development of the different
categories. It would require a map, diagram, or model in order to
show the parallel development of the subtribes Balsamocitrinae and
Citrinae. Here in the printed text where the subtribes are arranged
in the linear order of the pages and lines of the printed book, the
Balsamocitrinae fall at the end of the series, although in reality the Citrinae
are more highly developed with their extraordinary pulp-vesicles that fill all
the space not occupied by the seeds in the locules of the fruit.
However, to intercalate the subtribe Balsamocitrinae between the subtribes
Triphasiinae and Citrinae would obscure the close relationship clearly
exhibited by some of the genera of the subtribe Triphasiinae with some of the
more primitive genera of the tribe Citrinae. Keys to the groups and the genera of the subtribe Balsamocitrinae
are presented.
GROUP A. THE TABOG
GROUP
This group contains only one genus,
Swinglea, the tabog of Luzon, the northernmost island of the Philippine
Archipelago. It stands alone in having an oval,
longitudinally-ribbed fruit with a tough, leathery shell, not a hard, woody
shell as in all the other genera of the subtribe. Then, too, the
locule walls are lined with large mucilage glands that probably discharge into
the seed cavity the mucilaginous material in which the seeds are
embedded. No such mucilage glands have yet been found in other
genera of the Balsamocitrinae, but they may occur in less striking form in the
thick locule walls of Aegle and possibly other genera of this
subtribe. The seeds of Swinglea glutinosa are densely woolly,
much like those of the Indian bael fruit, Aegle marmelos. The
two genera, however, differ greatly in other important taxonomic characters.29
All species of Citrus so far tested graft
readily on young rapidly growing seedlings of tabog (Swinglea) used as a
rootstock and reach large size if planted in soil that stays warm the year
round, as for instances greenhouses. Citrus does not make a
vigorous growth when grafted on any other genus belonging to the subtribe
Balsamocitrinae. This would seem to indicate that Swinglea
is more closely related to Citrus than are the other members of the
subtribe.
XXVII. Swinglea Merrill
XXVII.
Swinglea Merrill, Jour. Arnold Arbor. 8:131. 1927.
Chaetospermum (Roem.) Swing. Jour. Wash. Acad. Sci. 3:101. 1913,
non Saccardo, Syl. Fungor. 10:706. 1892; Limonia subg.
Chaetospermum Roem. Syn. Hesper. 1:39. 1846. Illus. Tillson
& Bamford, Amer. Jour. Bot. 25:786, fig. 46. 1938; fig. 3-51 this work.
Type
species.—Limonia glutinosa Blanco = Swinglea glutinosa (Bl.)
Merr.
Distribution.—Philippines: Luzon
Island.
Common name.—Philippine tabog
or swinglea.
In 1913, Swingle published a
description of this genus under the name Chaetospermum as follows: "A
genus related to Belou [= Aegle], from which it differs in having
persistent leaves with small rounded sessile lateral leaflets, fewer stamens
(twice as many as the petals), fewer ovarial locules (8 to 10), an oblong
ribbed fruit with a thick leathery rind and cells [locules] lined with a spongy
tissue containing many large cavities or vacuoles.
"Leaves persistent, trifoliate, lateral
leaflets small, sessile, usually less than one-third as long as the median,
more or less blunt at the base or even rounded. Terminal leaflet
gradually narrowed at the base. Petioles narrowly winged with
a joint at the point of attachment of the leaflets. Spines
slender, straight, sharp, in pairs at the axils or else one of the spines is
replaced by a branch. Inflorescences axillary, composed of
from one to several flowers on rather long, slender pedicels.
Flowers perfect, 5-merous; calyx 5-lobed, petals 5, stamens 10,
free. Pistil with a well developed style and a thick rounded
stigma. Ovary with 8 to 10 cells [locules], each containing
numerous ovules. Fruit oblong, longitudinally ribbed, with a
very thick leathery rind, and with cells [locules] (filled with gum ?)
surrounded with watery tissue containing large cavities or
vacuoles. Seeds numerous in the long narrow cells, flattened
ovate, hairy. Germination—cotyledons aerial, not increasing
in size; first foliage leaves opposite, broadly ovate, subseriate, sessile,
abruptly narrowed at base." (Fig. 3-51.)
This genus differs
rather widely from the other genera of the Hard-Shelled Citroid Fruit Trees
(subtribe Balsamocitrinae) in having ellipsoid, or ovoid, ribbed fruits with a
leathery outer shell showing very long, pointed, radially arranged oil glands
in the peel and thick tissue containing mucilage glands lining the radial walls
of the segments surrounding the seeds. Its paleate seeds resemble
somewhat the woolly seeds of Aegle, and its trifoliolate leaves have a
general similarity to those of Aegle, Balsamocitrus, and
Afraegle, but its fruits differ widely in structure from those of other
members of the subtribe.
Swinglea makes
a good rootstock for Citrus if planted in soil that is always warm; it is
a much better rootstock for Citrus than any other Hard-Shelled Citroid
Fruit Tree that has been tested so far. Another indication of the
rather close relationship of Swinglea to Citrus and the five other
closely related genera constituting the True Citrus Fruit Trees is that it is
the only genus outside of this group that is attacked in the wild state by
citrus canker, a disease caused by the bacterial parasite Pseudomonas
citri. Lee (1918, p. 664) stated concerning Swinglea
glutinosa, which grows naturally and is also cultivated at the Lamao
Experiment Station near Manila, P.I., not far from a large collection of
Citrus species and cultivated varieties: "The susceptibility of C.
[haetospermum = Swinglea] glutinosa to canker is easily
greater than that of the sweet orange (Citrus sinensis) in the
Philippines." It should be remarked that no natural infections were
observed on any of the many other Citrus relatives under observation with
respect to their susceptibility to citrus canker, although some of them showed
slight susceptibly when inoculated by the needle-prick method (see Lee, 1918;
Peltier and Frederich, 1920).
It must be
remembered that the true taxonomic position of Swinglea is still somewhat
in doubt and that it may prove to be rather closely related to the Primitive
Citrus Fruit Trees, of which Hesperethusa and one species of
Pleiospermium also show a certain degree of relationship to Citrus
by being attacked, in the wild state, by a fungus belonging to the genus
Elsinöe, closely related to E. fawcettii Jenkins that causes scab
or verrucosis on several species of Citrus.
Burkillanthus, another of the Primitive
Citrus Fruit Trees, of the subtribe Citrinae, shows striking similarities to
Swinglea in its large fruits with stiff leathery rinds, its frequently
trifoliolate leaves, and in the size, shape, and structure of its flower parts;
but it differs widely from Swinglea in the anatomy of its fruits and in
its seeds. It would be very desirable to make comparative studies of
the development of the fruits of Burkillanthus and Swinglea and to
make grafting tests in order to determine the degree of relationship of these
two curious and little-known genera.
Swinglea glutinosa (Blanco) Merr. Jour. Arnold Arbor. 8:131. 1927.
Limonia glutinosa Blanco, Fl. Filip. 358. 1837; Feronia ternata
Blanco, Fl. Filip. 2:252. 1845; Aegle decandra Naves apud
F.-Villar, Nov. App. in Blanco, Fl. Filip. 38. 1880; A. glutinosa (Bl.)
Merr. Phil. Gov. Lab. Bur. Bul. 6:12. 1904; Limonia engleriana
Perk. 1905; Belou glutinosa (Bl.) Skeels, U.S. Dept. Agr. Bur. Pl. Ind.
Bul. 162:26. 1909; Chaetospermum glutinosum Swing. Jour. Wash.
Acad. Sci. 3:102. 1913. Illus. Naves, in Blanco, Fl. Filip.
2:pl. 124. 1880; Vidal & Soler, Sinop. Fam. Gen. Pl. Leñosas Filip.
Atl. pl. 25, fig. J(1-5). 1883; Swingle, Bul. Soc. Bot. France
58(mém. 8d):pl. 5. 1912; fig. 3-51 this work.
Type.—Wanting. Substitute type: (Merrill, Plantae
Blancoanae, No. 607). Herb. Bur. Sci., Manila.
Distribution.—Philippines: Luzon
Island.
Common name.—Tabog or
swinglea. Native name: Tabog or boyag (in Tagalog language,
Philippines).
Blanco's original description of
this plant follows (translated from the Spanish): Trunk with large solitary
spines. Leaves alternate, ternate. Leaflets
lanceolate, emarginate, shining, the central one largest. Calyx
with 5 teeth. Corolla with 5 thick linear petals, much longer than
the calyx. Stamens 10. Ovary cylindrical.
Style and stigma thick. Fruit oblong (3 in. [7.6 cm] long and more
than 2 in. [5 cm] thick) with protuberances and confused furrows on the
surface, with about 10 locules and some seeds in each, obovate compressed,
ending in a tuft of wool. Tree 8-10 yards high, [trunk] not very
thick. It is used for pillars in houses. The fruit is
aromatic and the juice sour. It is like a lemon and is used for
'cola.' I saw it on Mt. Arayat. Flowers in
Dec. [Spanish name] Mala cabuyao, Phil. name Tabog."
This original description may be supplemented
from herbarium material from Luzon Island, P.I., and from living plants in the
greenhouses of the U.S. Department of Agriculture as follows: Small or
medium-sized tree; twigs angled when young and finely pubescent, soon becoming
terete and losing pubescence; spines finely pubescent, 1 or 2 in the axils of
the leaves, especially on young vigorous shoots (flowering twigs frequently
spineless); leaves 3-foliolate, terminal leaflets 8-12 X 3-5 cm, oblanceolate,
bluntly pointed or sometimes rounded at the apex, gradually narrowed into an
acuminate, finely pubescent base which is not, however, differentiated into a
petiolule, although winged for 1/2-1 cm and articulated by means of a
pulvinus-like base, lateral leaflets oval or obovate, 2.5-5 X 1.2-2.5 cm,
1/5-1/2 the length of the terminal one (never more than half as long as the
terminal one!), bluntly cuneate and pubescent at base with a very short
pulvinoid petiolule, 1-1.5 mm long, not articulated with the blade, but
distinctly articulated with the petiole which is extraordinarily variable in
length (0.5-5 cm), flattened above, finely pubescent, very narrowly winged;
flowers solitary or in clusters in the axils of the leaves, or sometimes
terminal on short twigs leafy at the base; buds long, elliptical, 6-7 X 3-3.5
mm; calyx 2.5-3.5 X 1.5-2 mm, cup-shaped, with 5 blunt lobes; corolla white,
petals 5, 12-14 X 3-3.5 mm (in dried specimens); stamens 10, filaments
filiform, glabrous, anthers linear, 3.5-4 X 1 mm; pistil 7.5-9 mm long, ovary
short-stalked, obovate, pilose with stiff, yellowish-gray, bristling hairs,
style slender, 0.6-0.7 mm diam., sparingly hairy (hairs like those on the
ovary), twice as long as the ovary, longitudinally furrowed (showing 8-10
furrows), stigma 1.3-1.6 mm diam., flattened like a mushroom, apparently
faintly 10-furrowed; young fruit showing the persistent style; mature fruit
oblong-ovoid, 5-10 X 3-6 cm, locules 8-10; segment wall thickened, containing
numerous large, elliptical mucilage glands; seeds woolly, several in each
locule, surrounded by glutinous pulp; peel leathery with long, pointed, radially
arranged oil glands.
The fruit of
Swinglea shows remarkable structures, here called tentatively mucilage
glands, that seem to arise lysigenously, just as do the oil glands of the peel,
leaves, et cetera. These ellipsoid lysigenous mucilage glands
arise rather close together in the thick locule walls, both dorsal and
radial. Those on the outer portions of the radial walls slant
obliquely away from the middle of the locule walls to the membrane that lines
the narrow seed cavity at an angle of about 45° to the radial
walls. Consequently the mucilage glands in the radial walls of
adjoining locules are disposed at right angles to one another in a sort of
herringbone pattern; at any rate they are thus disposed in young fruits when
all the glands in the locule walls are nearly equal in size before the growing
seeds have more or less displaced and distorted the glands nearest the axis of
the fruit.
It is not impossible that similar,
less evident, structures will be found in the thick radial locule walls of
Aegle. The thin (2 mm) radial locule walls of
Aeglopsis show numerous but very different glands that secrete mucilage,
as described by Chevalier in Swingle (1912b, p. 241, pl. 3, fig.
3). At any rate the mucilage glands of Swinglea are of such
large size (5 to 10 by 2 to 3.5 mm) that they cannot be overlooked (see fig. 3-51).
The floral anatomy
of Swinglea was found by Tillson (1938, pp. 22, 30, fig. 46; also Tillson
and Bamford, 1938, pp. 789, 790) to be markedly different from that of the
other genera of the Hard-Shelled Citroid Fruit Trees of the subtribe
Balsamocitrinae in that the lateral sepal bundles are fused as they leave the
axis with the petal midrib bundles. This remarkable condition has
also been found in the five genera of the Primitive Citrus Fruit Trees, in the
species of Citrus belonging to the subgenus Citrus, and in
Paramignya and Luvunga, all belonging to the tribe Citreae.
The tabog, as this species is called in the
Philippines, is a small thorny tree widely distributed in Luzon
Island. It was found impossible to top-graft Citrus scions
on tabog trees growing out of doors at Manila, P.I., but in the Washington,
D.C., greenhouses of the U.S. Department of Agriculture vigorous young seedling
tabog plants took Citrus buds very well, and these buds continued to grow
vigorously and to flower and fruit freely for many years.
From the extraordinary energy of root growth of
the tabog observed in greenhouse sections kept at very high temperatures (up to
32.2° C, or 90° F), it seems probable that the tabog requires higher soil
temperature during the winter than is found in the citrus-growing regions of the
mainland of the United States except in extreme southeastern Florida, where the
tabog has made a very good growth in the sandy-loam and porous-limestone soil
characteristic of the Miami region and has withstood temperatures as low a 35° F
without loss of its leaves. In the Panama Canal Zone in the
Experiment Gardens at Summit, the tabog was reported by J. E. Higgins to make a
strong tree that fruited freely and yielded abundant viable seeds.
Since World War I, the tabog has been introduced to many countries in tropical
Central and South America, where it grows vigorously at altitudes of 1,500
meters or less (fig. 3-52).
This species should
certainly be given serious consideration as a rootstock for Citrus to be
grown in greenhouses where the temperature is warm all winter, and it should
also be tested in tropical regions having high soil temperatures throughout the
year.
GROUP B. THE BAEL-FRUIT
GROUP
The genera of this group
(Aegle, Afraegle, Aeglopsis, and Balsamocitrus)
represent the typical Hard-Shelled Citroid Fruit Trees and are obviously
closely related to one another in spite of their discontinuous distribution,
Aegle being limited to southeastern Asia and neighboring East Indian
islands and the other three genera to tropical and warm subtropical
Africa. All four genera have rather large fruits varying from the
size of a small orange (Aeglopsis) to the size of a small pummelo
(Aegle and Balsamocitrus). All the genera of group B
have fruits with a dense, hard, woody rind without longitudinal ridges like
those found in the leathery rind of the tabog (Swinglea) of group A, and
also without any sign of radially arranged shoe-peg-like elements, such as
constitute the woody rind of Feroniella.
Although all four of the genera that constitute
the Bael-Fruit Group have fruits resembling those of Citrus in size, in
the number of follicles, and in the number and arrangement of the seeds, none
show any sign of the highly developed pulp-vesicles that characterize
Citrus and the other genera of the True Citrus Fruit Trees.
None of the Bael-Fruit Group will hybridize with Citrus.
However, they have been successfully grafted on Citrus species under
greenhouse conditions at the Citrus Research Center in Riverside, California.
XXVIII. Aegle Corrêa
XXVIII.
Aegle Corrêa, Trans. Linn. Soc. 5:222. 1800. Belou Adans. Fam.
2:408. 1763.
Type
species.—Crataeva marmelos L. = Aegle marmelos (L.)
Corrêa.
Distribution.—India: wild in
large colonies in northern India up to 1,219 m, also in central and southern
India and Burma; widely cultivated in southeastern Asia and in the East Indian
Archipelago.
Common name.—Indian bael
fruit.
A thorny tree with deciduous, dimorphic
3-foliolate leaves; leaflets crenate, articulated with the petiole, thin and of
soft texture, with numerous translucent oil glands; inflorescences axillary,
loosely branched; pedicels shorter than the petals; flowers large; calyx
deciduous, small, saucer-shaped, with 4-5 short, blunt lobes; petals 4-5,
large, imbricate, long-ovate, with bluntly rounded apices; stamens very
numerous, with subulate filaments and long, linear anthers; disk small,
annular; ovary cylindric, with 8-20 locules, each containing numerous ovules in
2 rows; ovary tapering slightly toward the top and somewhat narrowed into the
style, which is about 1/4 or 1/5 as long as the ovary; stigma with longitudinal
furrows, cylindrical or bluntly conical, slightly wider than the style; fruits
globose or subglobose with a very hard, woody shell, segments 8-16 or more
(Talbot, 1909, p. 205, stated: "8-20"), narrow, filled with 6-10 or more seeds
imbeded in transparent glutinous gum, lateral walls of the segments thickened
and fleshy; seeds oblong, slightly flattened, woolly, monoembryonic; cotyledons
hypogeous in germination.
The genus
Aegle is one of the three monotypic genera of the orange subfamily
(Poncirus and Feronia are the other two) with deciduous
leaves. Two other genera of the subfamily, Clausena and
Murraya, have deciduous-leaved species: Clausena has three species
(C. pentaphylla, C. dentata var. dulcis, and C.
suffruticosa) and Murraya has one species (M. alternans).
Aegle is obviously related to the genera
Balsamocitrus, Afraegle, and Aeglopsis of tropical Africa
but differs from all of them in having very numerous (30 to 40 or more) stamens
(6 to 8 times as many as the petals) and very numerous (8 to 16 or more) locules
in the ovary, and also in having woolly seeds. It is a highly
specialized genus having a certain degree of resistance to cold.
Aegle marmelos (L.) Corr. Trans. Linn.
Soc. 5:223. 1800. Crataeva marmelos L. Sp. Pl. 444. 1753; Belou
marmelos (L.) Lyons, Plant Names 69. 1907. Illus. Roxburgh, Pl.
Corom. 2:pl. 143. 1798; Wight, Icon. Pl. Ind. Or. 1:pl. 16. 1840;
Bentley & Trimen, Med. Pl. 1:55. 1880; Wilkins, Hindu Myth. 390.
1882; Bonavia, Cult. Orang. Lem. Ind. Cey. Atl. pls. 242, 243. 1890; Beddome,
Fl. Sylv. 1:pl. 121. 1869; Turner, Agric. Gaz. N. S. Wales 4:pl.
14. 1893; fig. 3-53 this work.
Type.—Ceylon [cult.] (Hermann). Herb. Brit. Mus., London.
Distribution.—India, Burma, Thailand (?),
Indo-China (?); widely cultivated in southeastern Asia and the East Indian
Archipelago; native in northern India, often growing in large groups, up to
1,219 m altitude (4,000 ft); also native in central and southern India and
Burma.
Common name.—Indian bael
fruit.
Bentley and Trimen (l.c.) gave the
following description of this species: "A tree reaching a height of 30 or 40
feet when cultivated, with a short thick trunk and narrow oval head; in the
wild state smaller and more irregular, with short, strong, sharp, spiny
branches 1 inch or more in length in the axils of the leaves; bark bluish-grey,
soft, with irregular furrows on the younger branches. Leaves
alternate, compound, with one (rarely 2) pairs of shortly stalked opposite
leaflets, and a larger long-petioled terminal one, leaflets 1-2 inches long,
ovate or oval-ovate, abrupt or tapered at the base, somewhat attenuated towards
the blunt apex, very shallowly serratocrenate, smooth, thin, midrib prominent
beneath. Flowers 3/4 inch wide, sweet-scented, stalked, solitary,
or in few-flowered, lax, erect, axillary or terminal cymes. Calyx
shallow, with 5 short, broad teeth, pubescent outside. Petals 5
(rarely 4), oblong-oval, blunt, thick, pale greenish-white, dotted with glands,
imbricate, spreading. Stamens numerous, sometimes coherent in
bindles, hypogynous with short filaments half as long as the linear
anthers. Disk none or very small. Ovary oblong-ovoid,
slightly tapering into the thick short style which is again somewhat thickened
upward, stigma capitate, axis of ovary wide, cells numerous, 8-20, small,
arranged in a circle, with numerous ovules in each cell. Fruit
usually globose, 2-5 inches in diameter, pericarp nearly smooth, greyish-yellow,
about 1/8 inch thick, hard, filled with softer tissue becoming very hard and
orange-red when dry; cells as in ovary. Seeds very numerous,
somewhat compressed, ranged in closely packed tiers in the cells and surrounded
by a very tenacious, slimy transparent mucus which becomes hard when dry; testa
white, covered with woolly hairs immersed in the mucus, embryo with large
cotyledons, and a short superior radicle; no endosperm."
Aegle marmelos has dimorphic twigs:
(a) normal twigs with internodes 3 to 5 cm long with one well-developed
leaf at each node, often with one or two spines alongside; (b) foliage
spurs produced on primary branches of the previous year's growth, usually very
short, 1 to 3 cm long, with numerous very short internodes, each node bearing a
leaf but no spines. The numerous leaves crowded on the foliage
spurs vary greatly in size, the largest being nearly as large as the normal
leaves on rapidly growing long-internoded branches but having decidedly longer
petioles. The smaller leaves borne near the base of the foliage
spurs are often much dwarfed, sometimes being only one-fifth or one-tenth as
long as normal leaves. These crowded leaves of all sizes often hide
almost completely the branches which bear them.
Because of these dimorphic characteristics the
bael-fruit tree presents a peculiar appearance, with its long, slender young
branches with only a few leaves scattered along them arising from an inner crown
of older branches almost completely covered with massed foliage borne on the
leaf spurs. Poncirus trifoliata has somewhat similar dimorphic branches and
leaves but shows much less variation in the size of the leaves produced on the
leaf spurs.
Besides this dimorphism of twigs
and leaves, Aegle marmelos shows great variability in both kinds of
leaves on different seedling trees, not only in size but in important botanical
characters, such as the presence or absence of a separative layer at the
junction of the terminal leaflet with its petiolule, the relative length of
this petiolule and of the petiole, as well as the length of the petiole in
relation to the length of the entire leaf. The petioles on some
trees have distinct wings on each side for almost their entire length and on
other trees show only two inconspicuous green lines broadened into very narrow
wings at the upper end of the petiole. The leaflets vary greatly on
different seedling trees in marginal crenulation and flatness or curvature of
their surface. There is also great variation in the posture of the
leaves on the twigs which bear them and the degree to which the blades,
petioles, and supporting twigs show reddish coloration where exposed to
sunlight.
These surprising diversities in leaf
characters were studied by Swingle in October, 1941, on some thirteen fruiting
trees growing at Coconut Grove and Homestead, Florida, and convinced him that
it would be necessary to make a detailed study of both wild and cultivated
bael-fruit trees in India. The extraordinary variation in
taxonomically significant characters shown by the trees grown from seeds
imported from India very probably means that there are several different
strains, botanical varieties, or even good species to be found among the wild
plants growing in the mountains of northern India. Seedlings of
these diverse wild forms planted together in the villages would doubtless be
cross-pollinated by insects and produce complex and highly variable hybrids
like those we find growing in Florida.
For
many years few or no fruits of Aegle marmelos were produced by trees
growing in the continental United States, but now the trees in southern Florida
have fruited freely. S. J. Lynch, former horticulturist of the
Subtropical Branch Station of the Florida Agricultural Experiment Station at
Homestead, found that many bael-fruit trees in southern Florida were suffering
from zinc deficiency. Application of small amounts of zinc sulphate
caused them to make a vigorous new growth with green (not yellowish) leaves
and favored the setting and maturing of a good crop of fruit.
The Hindus esteem the ripe bael fruit very much;
in fact, many of them consider it to be the best of the citrus
fruits. Some European residents of India have been known to develop
a decided liking for it. Watt stated (1889, vol. 1, p. 123): "The
fruit, when ripe, is sweetish, wholesome, nutritious, and very palatable, and
eaten by all classes. The ripe fruit diluted with water, forms with
the addition of a small quantity of tamarinds and sugar, a delicious and
cooling drink." Concerning its medical value, W. R. Mustoe, then
superintendent of the Government Archipelago Gardens, Lahore, India, wrote to
David Fairchild on December 3, 1908, as follows: "All Indian medical
authorities agree that the Bael fruit has a most salutory influence on the
digestive system. The ripe fruit is mildly laxative and is a good
simple remedy for dyspepsia. The unripe fruit is a specific of the
highest value for dysentery so mild that it can be given to children without
danger." Roxburgh stated (1795-1819, vol. 2, p. 23, pl. 143): "The
fruit, delicious to the taste and exquisitely fragrant, is not only nutritious,
but possesses a laxative and aperient quality, confirmed by experience, which
renders it particularly serviceable in habitual costiveness."
The wild forms of Aegle marmelos have
small fruits (5 to 7.5 cm in diameter), whereas in cultivated forms the fruits
may attain a diameter of 12.5 to 17.5 cm (see fig. 3-53). According to Davis (1930, p. 111), in its
wild or semi-wild state the bael-fruit tree grows freely in the Bahraich Forest
in northern India (one of the northern districts of Oudh, in Uttar Pradesh
State to the south of Nepal), even on poor clay soils where other trees
fail. In view of the widespread use of the fruit in India both for
food and for medicine, it is very desirable that this species be tested in the
United States. Fortunately it is easily propagated from seed and
from layers. According to Turner (1893, p. 152), A. marmelos
has been successfully fruited in northeastern New South Wales as far south as
the Clarence River (about Lat. 29° 30' S.). It has for some time
fruited in the warmer parts of the United States, and it is almost certain to
succeed in Puerto Rico and Hawaii. In extreme southeastern Florida
this species has been grafted successfully on Aeglopsis chevalieri and on
Afraegle gabonensis, both of which are rather closely related plants that
grow vigorously in the sandy-loam, porous-limestone soils found in this
region.
Although the bael-fruit tree grows
commonly in tropical climates, it loses its leaves in winter when in a cool
climate; moreover, it is able to endure low temperatures in India when in a
leafless condition—as low, it is said, as 17.5° F (-8° C). It is
possible that, like many other deciduous trees, it must be exposed to the proper
degree of cold over a long enough period for food materials stored in the trunk
and twigs to be rendered available to support the new growth in early spring.
XXIX. Afraegle (Swing.) Engler
XXIX. Afraegle (Swing.) Engler, Die Pflanzenwelt Afrikas
3:761. 1915. Balsamocitrus Sec. Afraegle Swing. Bul. Soc.
Bot. France 58(mém. 8d):233. 1912.
Type species.—Citrus paniculata
Schum. = Afraegle paniculata (Schum.) Engl.
Distribution.—West Africa: Ghana, Togo,
Dahomey, Nigeria, Cameroon, Gabon, Fernando Po Island.
Common name.—African powder-flask
fruits.
Swingle described Afraegle as a
section of Balsamocitrus in 1912 and at the same time established the
section Eubalsamocitrus for the typical species, B.
dawei. The section Afraegle was raised to generic rank in
1915 by Engler. In his revision of the whole plant family Rutaceae,
Engler described it (1931, p. 352) as follows (in translation): "Calyx small,
5-merous, persistent. Petals 5 linear, imbricate.
Stamens about 20 with subulate filaments and anthers of about equal length,
inserted in a broad lobed disk. Ovary short-ovate, roughened with
glands, with about 8 locules, more or less, with numerous ovules in each
locule; style shorter than the ovary with a long-ovate stigma.
Fruits large, globose or pear-shaped, with a hardish shell,
many-seeded. Seeds large, up to 1.7 cm long, 1 cm thick, ovate, or
wedge-shaped at the base, rounded or bluntly angled. Trees with
trifoliolate, long-slender leaflets narrowed toward both ends, with axillary
thorns and small 6-10-flowered racemes."
This
West African genus of Hard-Shelled Citroid Fruit Trees contains three or four
species of medium- or large-sized trees, including A. (?) asso,
the tallest African species of the orange subfamily. Afraegle
is more advanced from an evolutionary point of view than Balsamocitrus
in that it has stamens three to four times as numerous as the petals
(instead of only twice as many). Afraegle is related to
Aeglopsis; the latter, however, has simple leaves and only six locules in
the ovary, instead of eight to ten, as in Afraegle. A key to
the species of Afraegle is presented.
1. Afraegle paniculata (Schum.) Engl. Die Pflanzenwelt
Afrikas 3:761. 1915. Citrus paniculata Schum. Dan. Vid. Selsk.
Afh. 4:153. 1827; Aegle barteri Hook. f. 1895; Limonia
warneckei Engl. 1905; Balsamocitrus paniculata (Schum.) Swing. Bul.
Soc. Bot. France 58(mém. 8d):231. 1912. Illus. Hooker f.,
Icon. pl. 2285. 1895; Swingle, loc. cit. pls. 1, 2. 1912; idem, in
Bailey, Stand. Cycl. Hort. 1:444, fig. 455. 1914; Engler, loc.
cit. 1915; idem, in Die Nat. Pflanzenfam. 19a:351, fig. 161.
1931; Tillson & Bamford, Amer. Jour. Bot. 25:786, figs. 39-45. 1938;
fig.
3-54 this work.
Type.—West Africa,
Gold Coast (Thonning, No. 179 ?). Herb. Copenhagen (?).
Cotype: Herb. Jussieu, Mus. Hist. Nat., Paris.
Distribution.—West Africa: Liberia (?),
Ivory Coast, Ghana, Dahomey, Nigeria.
Common name.—Nigerian powder-flask fruit.
In 1912 Swingle quoted a detailed description of
this plant by Chevalier. It reads as follows (in translation from
the French): "Tree 8-15 m high, branching 1.5-2 m above ground.
Trunk 25-40 cm diam., bark brown, lightly split longitudinally.
Branches numerous, spiny, making a rounded head like an orange
tree. Young twigs green, slender, glabrous, spines straight, very
sharp, 12-18 mm long, occurring in the axils of some leaves.
"Leaves alternate, 3-foliolate or, by exception,
1-foliolate by abortion, 8-16 cm long, completely glabrous.
Leaflets similar, oboval or oblong, cuneiform at base, rounded or alternate,
subacute at the apex, sometimes retuse, lightly crenulate in the upper half or
2/3 of the margin, firm or subcoriaceous, a beautiful green color both above
and below, central leaflet a little larger than the lateral ones, measuring
4.5-11 cm long and 2.5-5.5 cm broad, borne on a petiole 5-30 mm long,
articulated at the two ends, lateral petioles 2-10 mm long.
Inflorescences small, axillary panicles, 4-6 cm long, few-flowered (6-10 fls.),
isolated at the axil of the leaves or inserted on parts having already lost
their leaves. Rachis and pedicels glabrous green, pedicels 2-3 mm
long. Flowers white, with a strong odor of 'berlingot,' glabrous,
buds ovoid. Calyx cyathiform, 3-4 mm long with 4 short lobes
rounded and irregular. Petals 4, oval, oblong, rounded at the apex,
greenish outside, white inside, 10-13 X 4-6 mm, curved backward.
Stamens 15-20 (usually 16), filaments free, white, fusiform, erect, 4-5 mm
[long]; anthers oblong, 2.5 mm long. Disk greenish-stipulate,
slightly lobed. Ovary ovoid, greenish, divided into ±8
locules. Stigma ovoid, glandular, 3 mm long, borne on a very short
style.…
"Fruit globose or obovoid, sometimes
slightly stipulate at base, almost always depressed at the top, as large as a
big orange (6-8 cm diam. when mature), wrinkled on the surface, sometimes
tuberculous, without odorous glands.
"Pericarp
hard, 4-5 mm thick, resembling the pericarp of large fruited Strychnos,
remaining green when mature, then becoming grayish.
"Segments 8 (!) with smooth walls (not covered
with fleshy hairs). Each segment contains numerous seeds
irregularly arranged in 2 or 3 rows, inserted in the placenta, axillary and in
orderly arrangement, tegument whitish, parchment-like."
Fresh flowers of Afraegle paniculata from
a tree grown in the citrus greenhouse of the former Bureau of Plant Industry at
Washington, D.C. (see fig. 3-54), were compared on July 10, 1937, with fresh flowers of
A. gabonensis grown in the same greenhouse. They showed the
following points of difference: Inflorescences of A. paniculata much
longer, but pedicels slightly shorter (3 to 4 mm long); flowers about the same
size as those of A. gabonensis; calyx not flat and discoid but cup-shaped
with 3 to 5 clearly marked, very bluntly pointed lobes, finely ciliate at the
apex; petals very similar to those of A. gabonensis; stamens similar but
in A. paniculata nearly four times as numerous as the petals (often one
or two short of this number), whereas in A. gabonensis three times as
many (or a few short); disk smaller, about 3.5 to 4.5 mm wide and 1.5 mm high,
pea-green instead of yellowish-green, as in A. gabonensis.
This tree, which reaches a height of 15 meters,
is often planted by the natives in their villages in Dahomey and
Nigeria. The seeds are said to yield an edible oil and the leaves
are sometimes macerated and added to the water used for bathing. In
the United States, this species grows well in the greenhouses of the U.S.
Department of Agriculture and sometimes flowers, but so far has not borne
fruit. In the quasi-tropical coastal region near Miami, Florida,
which has an unusual type of soil consisting largely of porous-limestone rock
intermingled with fine sand or very sandy loam, this species, when well
fertilized, makes extraordinary growth—probably ten times as fast as in the
Washington greenhouse. A tree growing near Coconut Grove, when
only four or five years old and only 5 feet (1.6 to 1.8 m) high, had a lateral
spread of 15 to 20 feet (4.5 to 6 m). As it grew older, this tree
became much taller but still had long branches. Afraegle
paniculata grows in the Miami region much more rapidly than Aeglopsis
chevalieri (another vigorous but dwarfish West African Hard-Shelled Citroid
Fruit Tree), and is being tested as a rootstock for the bael fruit Aegle
marmelos, a species which often does not grow well on its own roots.
2. Afraegle gabonensis
(Swing.) Engl. Die Pflanzenwelt Afrikas 3:761. 1915. Balsamocitrus
gabonensis Swing. Bul. Soc. Bot. France 58(mém. 8d):233. 1912.
Illus. Swingle, loc. cit. 58(mém. 8d):235, fig.
B, pl. 4.
Type.—West Africa,
Gabon (Congo française) (R. Père Klaine, No. 2008); plant grown from seed of
this collection in greenhouse of Jardin des Plantes at Paris was for many
years grown in the citrus greenhouse of the former Bureau of Plant Industry,
Washington, D.C. (C.P.B. No. 7516). Natl. Herb., Washington,
D.C. Cotype: Gabon du Nord (R. Père Klaine, No. 1106, fruit
with seeds). Herb. Mus. Nat., Paris.
Distribution.—West Africa: Gabon.
Common name.—Gabon powder-flask fruit.
Swingle's original description of this species
reads as follows (in translation): "Balsamocitrus with simple or 3-foliolate
leaves, lateral leaflets much shorter than the terminal one (never half as
long), lanceolate or narrowly lanceolate, more or less acute at the apex, acute
at the deltoid base, margin almost entire or slightly crenate, densely
glandular-punctate; petioles winged, 0.5-3.5 cm long in 3-foliolate leaves,
much longer than in 1-foliolate ones, very glabrous, flattened above, spines
slender; sharp, solitary or in pairs in the axils of the leaves, scarcely as
long as the petioles, 1-2 cm long or very slightly recurved.
Flowers unknown. Fruits subpyriform, size of a large orange, cortex
hard, 3 mm thick; seeds numerous, glabrous, oval or cuneiform, more or less
angled, 1 1/2-1 3/4 cm long. 3/4-1 cm wide. Spiny tree,
young twigs virgate green."
Fresh flowers from
a tree grown in the former Bureau of Plant Industry greenhouse at Washington,
D.C. (tree grown from seed sent by R. Père Klaine [No. 2008] in 1892 from Gabon
to Paris, brought to the United States in 1912 by Swingle [l.c.]), were
examined. The inflorescences are short, axillary racemes or
corymbs, pedicels glabrous, 5-7 mm long, 1 mm diam. at base, 2 mm diam. at
junction with the calyx; calyx 3-4 mm diam., thick, flat, plate-shaped with
lobes very faintly or not at all marked, but with a thin, hyaline, ciliate
membrane around the margin (the interruptions in this membrane probably delimit
the poorly defined sepals); bud broadly conical, 4-5 mm wide and 7-8 mm long,
vivid green; petals 3-5, green without, greenish-white or cream-white where
thickest, 9-11 mm long, 3-5 mm wide, ovate-rounded at the tip and slightly
acute, curved, strongly incurved near the apex, soon falling, each carrying 2
stamens attached at the base; stamens apparently 3 times as many as the petals
(9-15), 4-6 mm long, glabrous, subulate, 1 mm diam. at the base, 0.5 mm at
attachment to anthers, which are linear, arising at the base of a very long
lobed disk and curving upward along the furrows in it; disk 5-6 mm diam., 2-3
mm high, forming a shallow cup in which the ovary fits loosely; ovary ovate,
about 3 mm high, 2 1/2-3 mm wide, with 8 locules; ovules in each locule
numerous, in 2 rows; style very short, thick, 1 3/4 mm wide, 1 mm long, ending
in an ovoid glutinous stigma, 2 mm long and 1 1/2 mm wide.
Afraegle gabonensis, like Aeglopsis
eggelingii, has three times as many stamens as petals,30 not twice as many or
four (or more) times as many as do the other species of the
subfamily. This species grows vigorously and fruits sparingly in
the Fairchild Gardens, Coconut Grove, Florida.
IMPERFECTLY KNOWN SPECIES
3. Afraegle asso Engl. Die Pflanzenwelt Afrikas 3:761.
1915.
Type.—Molunda district, southern
Cameroons, West Africa. Herb. Bot. Mus., Berlin-Dahlem.
Distribution.—Known only from the type
locality.
Engler's short description (l.c.)
reads in translation as follows: "Finally there is found in the District
Molunda, in South Kamerun, a tree up to 30 meters [98 1/2 ft.] high, with
broadly ovate leaflets and large fruits."
According to Engler, this species and A. milbraedii may belong to
Balsamocitrus instead of to Afraegle, as flowers (necessary to
decide to which genus they belong) are still unknown.
4. Afraegle milbraedii Engl.
Die Pflanzenwelt Afrikas 3:761. 1915.
Type.—San Carlos, Fernando Po Island, West Africa. Herb. Bot.
Mus., Berlin-Dahlem.
Distribution.—Known only from the type locality.
Engler gave (l.c.) a very brief notice of this
species as follows: "[Afraegle gabonensis] is rather closely approached
by A. Milbraedii from San Carlos on Fernando Po, of which unfortunately
we have only imperfect specimens; it is distinguished from A. gabonensis
by leaflets that are more linear than lanceolate but more strongly narrowed
into the petiolule at the base, and finely crenulate on the margins; the
pear-shaped, hard-shelled fruits are unfortunately not developed
normally." Since Engler (1931, p. 353), shortly before his death,
reprinted this unchanged, evidently no additional material had been obtained.
XXX. Aeglopsis Swingle
XXX.
Aeglopsis Swingle, Bul. Soc. Bot. France 58(mém. 8d):237.
1912.
Type species.—Aeglopsis
chevalieri Swing.
Distribution.—West Africa: Ivory Coast.
Common name.—Dwarf powder-flask fruit.
Much branched, very spiny small tree or shrub; leaves simple, subcoriaceous,
pellucid-punctate; petioles short, spines solitary, axillary; inflorescences
axillary panicles, few- or many-flowered; flowers 4-5-merous; stamens 2 or 3
times as many as the petals; style very short, stigma cylindric, viscous; disk
large, lobed, sulcate, annular, enclosing the ovary, which has 6-8 (rarely 5)
locules, ovules 12-18 (?) in each locule; fruit subglobose or pyriform, 5-11 cm
long, 4-8 cm diam., size of an orange, with a hard, woody shell, filled with
mucilaginous juice; seeds ovate-compressed, with a coriaceous, glabrous
testa.
This genus of two species, one found in
the Ivory Coast region of West Africa and the other one in Uganda and Sudan in
East Africa, is doubtless closely related to Afraegle,
Balsamocitrus, and Aegle. It is the simplest form of all
this group of Hard-Shelled Citroid Fruit Trees in that it has fewer locules in
the ovary (6 to 8 instead of 8 to 20); moreover, it has twice or three times as
many stamens as petals, a somewhat primitive character in the orange
subfamily. It shows, however, an advanced character in its
invariably simple leaves (even the first two opposite pairs of
post-cotyledonary leaves are simple). A key to the species of Aeglopsis appears below.
1. Aeglopsis chevalieri Swing.
Bul. Soc. Bot. France 58(mém. 8d):240. 1912. Illus.
Swingle, loc. cit. pl. 2, figs. 1-9, and pl. 3; idem, in Bailey,
Stand. Cycl. Hort. 1:223, fig. 130. 1914; fig. 3-55 this work.
Type.—Port de Sassandra, Ivory Coast, West Africa (Chevalier, No.
17940). Herb. Chevalier, Herb. Mus. Hist. Nat., Paris.
Paratypes: Herb. Mus. Nat., Paris, and Natl. Herb., Washington, D.C.
Distribution.—Collected only along the
Ivory Coast from western Sassandra to the Liberian frontier; doubtless occurs
also in Liberia and in Ghana.
Common
name.—Chevalier's aeglopsis.
Shrub or
small tree, branches numerous; spines solitary, straight, strong, pointed,
2-3.5 (sometimes 3-7) cm long; leaves simple, medium-sized, 5-12 X 2.5-7 cm,
elliptical, rounded at the apex or short-acuminate, cuneate at base and merging
gradually into the short, glabrous, wingless petiole (2-4 mm long), which is
not articulated with the leaf blade; inflorescences small, axillary panicles,
few-flowered (4-8) or sometimes many-flowered (20-40), pedicels slender (3-5 mm
long); flowers glabrous, white, calyx 4-5 mm diam., with 4-5 short, wide lobes
more or less rounded, petals 5 (sometimes 4), white, oblong-lanceolate, rounded
at the apex, 9-12 X 3.5-5 mm, caducous, stamens 8-10, filaments free, linear,
5-6 mm long, anthers oblong, 2.5-3 X 1 mm; disk subcupulate, very large, 5-6 mm
diam., 2.5-3 mm high, with about 10 rounded lobes; ovary ovoid, 2.5-3 X 2-3
mm, merging imperceptibly into the short style, locules 6 (sometimes 5),
containing numerous ovules (12-18 ?) arranged in 2 series; fruits globose or
pyriform, attenuate at the base when young, 6-9 cm diam., with 6 (sometimes 5)
segments, 8-12 seeds in each segment pressed together and imbedded in a liquid,
mucilaginous jelly secreted by the glands on the locule walls; peel
semicoriaceous, green, becoming yellowish, 3 mm thick, with many nonodorous
superficial glands; seeds ovoid, flattened, 12-16 mm long, 10-12 mm wide, and
4-7 mm thick, with a parchment-like, glabrous testa.
Aeglopsis chevalieri is the only
Hard-Shelled Citroid Fruit Tree that has flowered and set fruit freely in the
orange house of the U.S. Department of Agriculture at Washington, D.C., while
still a small tree, 2 or 3 meters high (fig. 3-55). It flowers profusely and bears a few
small, pyriform, only moderately hard-shelled fruits of brilliant orange-brown
color. The seeds germinate freely, but the cotyledons, although
turning green, remain buried in the soil. Seedlings of A.
chevalieri planted some years ago in David Fairchild's collection of
Citrus and related plants in his tropical garden, "The Kampong," at
Coconut Grove, Florida, have made a remarkably vigorous growth.
They have flowered profusely and have ripened abundant fruits filled with good
seeds.
Seedling plants of this species, so
easily obtained in quantity, make excellent rootstocks on which to graft
Balsamocitrus, Afraegle,and also Aegle, genera which seldom
fruit in the United States and which are also hard to propagate from
cuttings. Aeglopsis seedlings were used not only to propagate
the rare East African Balsamocitrus dawei by grafting the latter species
on the young seedlings but also to induce rooting of Y-cuttings of
Balsamocitrus dawei (see Swingle, Robinson, and May, 1929, p. 79).
2. Aeglopsis eggelingii M. R.
F. Taylor, Kew Bul. Misc. Inform. 1940:53. 1940.
Type.—Uganda, Bunyoro District (Lat. 1°
10'-2° 20' N., Long. 30° 40'-32° 25' E.), Siba Forest (Eggeling, No.
3006). Herb. Kew.
Distribution.—Uganda: between Lake Edward and Lake Albert, Toro District,
Bunyoro District; Sudan: Equatoria Province, Lado.
Common name.—Eggeling's aeglopsis.
The original description translated from Latin
reads: "Differs from A. Chevalieri Swingle in its shrubby habit, axillary
spines seldom paired, pedicels with bracts, globose buds, petals 3 or 4, ovate,
never lanceolate, stamens 3 times as many as the petals, ovary 7-8-locular,
Much-branched [sic] shrub, 4 m high, spiny, glabrous.
Branches rounded, compressed when young, brownish, internodes 2-4 cm long,
spines axillary, solitary or paired, 2-3 cm long, stiff, erect, spreading;
leaves simple, alternate, subcoriaceous; petioles 5-8 mm long,
canaliculate-pulvinate; leaf blade ovate or ovate-lanceolate, 7-14 cm long,
3-5.5 cm wide, apex obtuse or acute or often acuminate, cuneate towards the
base, margins slightly and irregularly crenate, dark green above, pale green
below, densely pellucid-punctate on both sides, midrib prominent, lateral veins
7-10 on each side, spreading, arcuate; inflorescence axillary, few-flowered
(about 5), peduncles irregularly branched, branchlets very short (up to 3 mm
long), pedicels 3 mm long 'ampliati' [swollen above ?], with 2 bracts,
glandular-punctate, bracts ovate-lanceolate, 1 mm long, obtuse; flowers 1.5-2.5
cm diam., greenish-white, buds globose or slightly 3-4-lobed, about 4 mm diam.;
petals 3-4, ovate, 1 cm long, 7-8 mm wide, obtuse at the apex, truncate at
base, thick, glandular-punctate, longitudinally striate above, margins
sometimes slightly incurved towards the apex; stamens 9-12 (3 times the number
of the petals), filaments strap-shaped, attenuate towards the apex, about 5 mm
long, 1 mm wide, compressed, thick, incurved; connective glandular at apex;
anthers oblong, 2 mm long, about 1 mm wide; disk annular, 5-6 mm diam., 1.5-2 mm
high, lobed, sulcate; ovary ovoid, 3-4 mm long, 2-3 mm diam., base sunk in the
disk, locules 7-8, ovules numerous; style 2 mm long, 1 mm diam., rounded;
stigma cylindrical, about 1.5 mm diam.; fruit subpyriform, 9-11 cm long, 7-8 cm
diam., fragrant, 7-locular, locules widely triangular, polyspermous, peel
subligneous; seeds subellipsoid, 1-1.5 cm long, 8 mm wide, compressed."
William J. Eggeling, who discovered this species
and in whose honor it was named, made an interesting report about it in a
general work on the forest trees of Uganda (1940, p. 205). He
stated that it is a spiny bush or tree, growing up to 20 feet high on the edges
of the forest; "spines strong, straight, sharp, slender 1/3-1/2 in. [8 to 13
mm] long"; also, "fruit yellow-green, woody, pear-shaped, up to 3 in. diam. [75
mm]; seeds surrounded by a strongly aromatic balsam."
XXXI.
Balsamocitrus Stapf
XXXI. Balsamocitrus Stapf,
Jour. Linn. Soc. 37:504. 1906.
Type
species.—Balsamocitrus dawei Stapf.
Distribution.—Uganda: a monotypic genus known only from the type locality
in the Budongo Forest, Unyoro.
Common
name.—Uganda powder-flask fruit.
Stapf's
original description of the genus reads, in translation, as follows: "Related to
Aegle, but distinct in habit of growth, in having thick leaves, and
chiefly in having few stamens and the seed with glabrous testa.
Flowers hermaphrodite. Calyx small, 5-merous, persistent for a
day. Petals 5, oblong, imbricate. Stamens 10; filaments
subulate, longer than the sagittate anthers, inserted [sic] in the
annular disk. Ovary ovoid, with 8 locules; style short,
conical-cylindric, soon deciduous; stigma inconspicuous, slightly sulcate;
ovules many in each locule, in 2 series. Fruit ovoid-globose, with
a woody cortex, 8 segments, filled with many seeds in a liquid balsamic jelly,
pulp scarce. Seeds subellipsoid, slightly compressed, testa
coriaceous, very glabrous. Endosperm lacking. Embryo
with large thick fleshy cotyledons, radicle small. Unarmed tree,
tall. Leaves 3-foliolate, leaflets thick, cortaceous, subcrenulate,
pellucid-punctate. Panicles axillary, short, flowers,
small. Fruit large. One species."
Balsamocitrus is apparently the most
primitive of the four closely related genera of Hard-Shelled Citroid Fruit Trees
(Balsamocitrus, Aeglopsis, Afraegle, and
Aegle). It is like Aeglopsis chevalieri in having only
twice as many stamens as petals but, unlike the simple-leaved Aeglopsis,
it retains the trifoliolate leaves undoubtedly ancestral in this group, and very
rarely produces unifoliolate leaves. Balsamocitrus does not
show the exuberantly-developed, lobed disk that is found in the other African
genera of this group, Aeglopsis and Afraegle, but has a simple,
annular disk like the Indian genus Aegle, which, however, differs from
all the African genera in having very numerous stamens and woolly seeds.
Balsamocitrus dawei Stapf, Jour. Linn.
Soc. 37:504. 1906. Illus. Stapf, loc. cit. pl. 37; Swingle,
Bul. Soc. Bot. France 58(mém. 8d):226, fig. A. 1912;
idem, in Bailey, Stand. Cycl. Hort. 1:443, fig. 454. 1914;
Swingle, Robinson, & May, Jour. Hered. 20(2):figs. 9-12. 1929;
Eggeling, Indig. Trees Ugand. fig. 58,a-d. 1940; fig. 3-56 this work.
Type.—East Africa, Uganda, Unyoro, in the Budongo Forest east of Lake
Albert Nyanza (Dawe, No. 788). Herb. Kew.
Distribution.—Known only from the type
locality.
Common name.—Uganda
powder-flask fruit.
The original description,
as translated, reads: "Tree, about 20 m tall, very glabrous. Twigs
rather thick, on drying black, young ones hollow or spongy
('fistulosus'). Leaflets unequal, ovate or ovate-elliptic,
somewhat obtuse, or subacuminate, acute at the base, crenulate, 4-7 cm long,
3.5-5 cm wide, on drying fuscous, black-punctate; lateral petiolules 7-10 mm
long, intermediate terminal ones up to 15 mm long; the common petioles 2.5-3 cm
long. Panicles 2-4 cm long, contracted; bracts very small; ultimate
pedicels up to 4 mm long, rather thick. Calyx up to 2 mm long,
segments broadly ovate, acute, somewhat thick. Petals white, 5 mm
long. Filaments 2 mm (or a little more) long; anthers scarcely 1.5
mm long. Fruit up to 13 cm long, 11 cm wide, cortex 5 mm
thick. Seeds up to 16 mm long, 10 mm wide, 8 mm thick.
Unyoro, Budongo Forest, Dawe 788."
Eggeling,
in his work on the forest trees of Uganda, stated (1940, p. 205): "Deciduous
forest tree, 70 ft.; flowers greenish-yellow in contracted axillary panicle,
2-3 in. [50-75 mm] long, petals 1/5 in. [5 mm] long [sic], very odorous,
woody (shell 1/5 in. [5 mm] thick); yellow-orange when ripe, ovoid globose, up
to 5 1/2 in. [13 cm] diam., 6-8 locules; seeds several per locule, about 2/3
in. [16 mm] long. Wood yellow-white, even in grain, hardy, nailing
badly but not splitting, difficult to saw, planing badly, turning well, taking a
good polish; weight 52 lbs. per cu. ft. air-dry."
This remarkable Citrus relative is native
to the plateau of Uganda, East Africa, to the east of Lake Albert, at altitudes
of 600 to 915 meters (2,000 to 3,000 ft.), where it attains a height of about 25
meters (82 feet). Seeds were collected by M. T. Dawe from the type
locality and sent to Swingle at the former Bureau of Plant Industry before
World War I. Under culture in the greenhouse it grew freely, but
even after many years did not flower. (See fig. 3-56.)
Balsamocitrus
dawei can be propagated easily by grafting on seedlings of Aeglopsis
chevalieri (which fruits freely in the greenhouse) and twigs can be forced
to root by the so-called nurse-grafted Y-cutting method (Swingle, Robinson, and
May, 1929, p. 79), where one fork of the Y is grafted on Aeglopsis until
the base of the Y strikes root. It is highly probable that
Balsamocitrus could be grafted also on the species of Afraegle and
on Aegle as well, as it is closely related to both of these
genera. As it is one of the largest trees among the Hard-Shelled
Citroid Fruit Trees (subtribe Balsamocitrinae), it should be tested as a stock
for Aegle, the bael fruit of India, the most important widely consumed
species in the subtribe but one that has proved difficult to grow on its own
roots in the United States.
GROUP C. THE WOOD-APPLE
GROUP
This well-marked group comprises two
genera, Feronia and Feroniella, which differ from all the other
genera of the subtribe Balsamocitrinae in having a most remarkable fusion of
the four to six locules of the ovary into a single cavity (showing when young
short rudiments of the follicule lateral walls that, however, do not reach to
the center of the ovary). As the core or axis of the ovary has
disappeared, an entirely new placentation has developed in this group which is
not found elsewhere in the orange subfamily or even elsewhere in the family
Rutaceae: a placenta develops along the middle of the ventral wall of each of
the original locules and bears very numerous ovules that because of their long
raphae are pendent and spread apart until they seem to cover the whole ventral
wall.
All in all, no other group of
Citrus relatives shows such striking deviation from the normal in ovary
and fruit structure. Nevertheless, Citrus can be grafted on
both Feronia and Feroniella, and there can be no doubt that the
genera of group C are related to those of group B. There is less
evidence of relationship of group C to group A, but plants of both of these
groups can be grafted on Citrus.
XXXII. Feronia Corrêa31
XXXII. Feronia Corrêa, Trans.
Linn. Soc. 5:224. 1800.
Type.—Feronia elephantum Corrêa = F. limonia (L.)
Swing.
Distribution.—India, Ceylon,
Burma, Thailand, Indo-China.
Common
name.—Indian wood apple.
Flowers in loose
panicles, hermaphrodite, by abortion polygamous; calyx small, 5-toothed,
caducous; petals 5, rarely 4-6, spreading, oblong or ovate-lanceolate,
imbricate in the bud; stamens 10-12; filaments dilated, bound together by
densely woolly pubescence at the base, subulate at the apex; anthers large,
linear-oblong, attached at the base, disk short; ovary globose, incompletely
4-6-locular, then 1-locular (with 4-6 parietal placentae), merging into a
short, thick, attenuate style; stigma oblong-fusiform; ovules at the internal
angles of the incomplete ovary walls, very numerous, in several series; fruit
large, globose, having a woody shell, with a single cavity, with numerous
parietal placentae, bearing numerous seeds surrounded by a gum-like pulp; seeds
oblong, compressed, testa thin, brown, hairy; cotyledons thick, fleshy, radicle
pointing away from the hilum. A medium-sized spiny tree with hard
wood. Spines short, straight, axillary. Twigs and
leaves densely covered with minute pubescence when very young, soon
glabrous. Leaves deciduous (in tropical climates often
persistent), odd-pinnate, leaflets opposite, subsessile, entire or slightly
crenulate, pellucid-punctate; petiole and rachis simple or winged.
This genus deviates widely from all the other
genera of the orange subfamily except the closely allied Feroniella in
having an ovary that in its earliest stages of development shows four to six
locules which soon fuse into a single locule with parietal placentae with very
numerous ovules. From Feroniella, to which it is obviously
closely related, it differs in having only twice as many stamens as petals
(instead of four times as many), stamens without basal appendages, hairy seeds,
and a fruit with a woody shell not composed of wedge-shaped radial
elements. The leaves of Feronia, as well as those of
Feroniella, are odd-pinnate with paired opposite leaflets on a rachis
that is composed of segments articulated at each leaf pair. This
character is found only in the tribe Citreae and there only in the
pinnate-leaved species. The fact that Citrus can be grafted
on Feronia is an indication of its not too remote relationship to the
True Citrus Fruit Trees.
Feronia
limonia (L.) Swing. Jour. Wash. Acad. Sci. 4:328. 1914. Schinus
limonia L. Sp. Pl. 1:389. 1753; Feronia elephantum Corr.
Trans. Linn. Soc. 5:225. 1800; Limonia acidissima L. 1762.
Illus. Roxburgh, Pl. Corom. 2:pl. 141. 1798; Beddome, Fl. Sylv.
pl. 121. 1871; Wight, Icon. Pl. Ind. Or. 1:pl. 15. 1840; idem,
Gartenflora 33:87. 1883, and 34:pl. 1206. 1885; Talbot, Forest Fl.
Bombay Presid. 1:204. 1909; Tillson & Bamford, Amer. Jour. Bot.
25:786, figs. 47-49. 1938; fig. 3-57 this work.
Type.—Ceylon (Hermann). Herb. Brit. Mus., London.
Distribution.—India, Ceylon, Burma,
Indo-China, Malay Peninsula, Indonesia.
Common name.—Indian wood apple.
This
species was described by Trimen (1893, p. 229) as follow: "A small tree,
branches numerous, with smooth, whitish bark and with sharp, straight, ascending
spines, 0.5-1.5 in. [1.2-3.8 cm] long; l. pinnate, 3-4 in. [7.5-10 cm long],
rachis and petiole flat, very narrowly winged, glabrous, lfts. opposite, in 2
or 3 pairs and usually a terminal one, nearly sessile, 1-1.5 in. [2.5-3.8 cm
long], oval or obovate, obtuse, entire, glabrous, basal ones the largest; fl.
small, numerous, in small, paniculate, sessile cymes from the axils of fallen
l., ped. slender, pubescent; cal. very small; pet. ovate, acute, spreading,
smooth; stam. 7-12, fil. very short, anth. very large; disk finely woolly; ov.
with numerous ovules in each cell, style very short, stigma fusiform; fruit
large, 2-2.5 in. [5-6.3 cm diam.], globose, hard, pericarp woody, rough,
whitish, 1-celled; seeds oblong, compressed." He stated further:
"Flowers Feb. Mar.; pale green, stained with red purple, anthers dark
red." Brandis (1906, p. 119) stated: "Flowers dull red, generally
unisexual, in lax panicles, male and bisexual flowers frequently on the same
inflorescence." He also noted that the leaves are
deciduous. (See fig. 3-57.)
In addition to the
description given above, it should be noted that the calyx is deciduous, as
Hooker (1875, vol. 1, p. 516) pointed out: "Calyx small, flat, 5-toothed,
deciduous." He also noted that the leaves smell of aniseed and are
deciduous. According to Guillaumin (1911, pp. 685, 687), the
anthers are "twice as long as the filaments."
Although apparently universally reported as glabrous throughout except for the
pedicels and peduncles, which are pubescent, and the base of the stamens, all
the specimens that Swingle examined (from southeastern Asia, as well as
greenhouse specimens from Washington, D.C.) show minute appressed
yellowish-gray or nearly colorless hairs covering the petiole, rachis, and
midribs of the very young leaves and also of the young twigs and
spines. The very youngest new growth is covered almost as
completely with these hairs as the very young growth of
Glycosmis. These hairs soon disappear except at the joints
of the rachis and base of the petiole. The seeds are pilose, as
noted by Corrêa in his original description of the genus Feronia.
The wood-apple tree (sometimes called elephant
apple), like the related bael-fruit tree (Aegle marmelos), also native in
the hill regions of northern India, is deciduous; however, it is not reported
as growing above 457 meters (1,500 feet) in the western Himalaya, whereas the
bael tree in the same region grows up to 1,219 meters (4,000 feet).
The wood apple is common in Ceylon in dry regions but the bael tree is not
native so far south. The wood-apple tree is often planted in
dooryard gardens in southern Asia and Java.
The ability to drop its leaves puts the wood apple in a special, small group of
Citrus relatives. It will be well worth while to study the
wood apple and the bael fruit as examples of subtropical Citrus relatives
that have acquired the deciduous habit.
As
Feronia limonia can be used successfully at least for a few years as a
rootstock for Citrus, it should be compared with the trifoliate orange
(Poncirus trifoliata), another deciduous Citrus relative that has
been used with great success as a rootstock for kumquats and for the satsuma
orange. Citrus when grafted on the wood apple is sometimes
forced promptly into bloom. Such a graft might be utilized in an
attempt to force newly originated or newly introduced varieties into bloom so
that they may be used promptly in making other desired hybrids.
The ripe fruits of the wood apple contain a
sour-sweet, aromatic pulp in which the seeds are imbedded. This
pulp is sometimes eaten mixed with palm sugar or sweetened and stirred into
coconut milk. It is also used to make a jelly said to be more or
less astringent. A chutney is made from the pulp that is much liked
by the natives. (See De Silva, 1890, p. 722; and Watt, 1890, vol.
3, pp. 326-27.)
XXXIII. Feroniella Swingle
XXXIII. Feroniella Swingle, Bul. Soc. Bot. France
59:776. 1912.
Type
species.—Feroniella oblata Swing.
Distribution.—Laos, Cambodia, North Vietnam, South Vietnam, eastern
Thailand, Java.
Common
name.—Feroniella.
A small tree, twigs
terete, with single axillary spines; leaves imparipinnate, with 3-6 or 7
strictly opposite pairs of leaflets, rachis segments and petiole cylindric, or
more or less broadly winged; leaflets small (1.5-4 X 0.8-1.5 cm), obovate or
oblong, rounded or bluntly pointed at the tip; inflorescences axillary,
panicles branched from the base, flowers perfect or male by abortion,
4-6-merous (commonly 5-merous), calyx small, with 4-6 acute lobes; petals 4-6,
lanceolate or oblong-lanceolate, very acute at the tip; stamens 16-20,
filaments slender, glabrous, each furnished at the base on the inner side with
a pilose linear appendage, free above, about 2/5 or 1/3 the length of the
filament, anthers oval, much shorter than the filaments; disk very short, ovary
subglobose, narrowed abruptly below and merging abruptly above with the slender
style as long or longer than the ovary, locules at first 5 or 6 but soon
confluent to 1 more or less lobed ovarial cavity with very numerous ovules on
parietal placentae; fruits large, globose or oblate, spheroidal, with a thick,
woody shell composed of radially arranged prisms; seeds oval, flattened, testa
membranaceous, glabrous; cotyledons epigeous in germination.
Feroniella has leaves much like those of
Feronia and also similar ovaries, with a single cavity formed by the
early confluence of the original five or six locules. It differs
from Feronia in having the stamens about four times as numerous as the
petals (instead of twice as many), in having the anthers much shorter than the
filaments, in having glabrous instead of pilose seeds, and especially in having
pilose, partly free, appendages at the base of the filaments. In
this last character it differs from all the other genera of the orange
subfamily. A key to the Feroniella species is presented.
All three species of Feroniella are
closely related to one another but the genus is not closely related to any genus
other than Feronia.
1.
Feroniella oblata Swing. Bul. Soc. Bot. France 59:779. 1912.
Feroniella lucida Guill. (non Scheffer), in Lecomte, Fl. Gén. Indo-Chine
1:684. 1911. Illus. Guillaumin, loc. cit. fig. 72(1-5);
Swingle, loc. sit. [sic] pl. 18; idem, in Bailey, Stand.
Cycl. Hort. 3:1219, figs. 1493, 1494. 1915.
Type.—French Indo-China, Cambodia,
Province of Samrong-Aong (L. Pierre, No. 652). Herb. Mus. Hist.
Nat., Paris.
Distribution.—Cambodia,
Laos, South Vietnam, eastern Thailand.
Common name.—Indo-China feroniella.
This species is a medium-sized spiny tree, 8-20 m high, native in the eastern
part of the Indo-Chinese Peninsula; it is characterized mainly by the leaves
with only 7-9 leaflets, which are oval-elliptic or obovate, 2-3 X 1-2 cm,
rounded at the apex or slightly emarginate, bluntly cuneate at the base, and
the cylindric, pilose petioles, 1-2.5 mm long. The rachis segments
are cylindric or narrowly winged. The flowers are grayish-white,
1.5-2.5 cm diam.; sepals very short, 1-2 X 0.5 mm, very pilose, soon falling;
petals lanceolate, 1.2-1.8 cm long, with very acute cuneate tips; ovary with
5-6 imperfect locules soon fusing into a single cavity. The
flattened-globose fruits, borne in clusters of 3 or 4, are 5.5-6.5 cm wide, 4.5
cm high, and have woody shells, 7-8 mm thick, composed of tapering prisms 1:5-3
mm diam. with the largest end out, and arranged like the stones of a curved
arch. The center of the fruit is filled with red flesh said to be
edible.
This species, rather common in the
forests of the plains and mountains of Indo-China, makes a medium-sized tree,
up to 20 meters in height, with brilliant, deep green foliage. The
fruits look like small green oranges. Swingle's citation of Lecomte
in the synonymy above and the illustrations cited for this species appear to be
incorrect. Guillamin [sic] (1911, fig. 72) presents an
obvious illustration of Feronia lucida instead of Feroniella
oblata. Since Swingle's original notes were not available for
clarification, the synonymy is retained intact. Further study will
be required to ascertain correct synonymy for Feroniella oblata.
Plants of this species grown in the U.S.
Department of Agriculture's greenhouses in the Washington, D.C. area were
extremely spiny and had small leaves, only 6 to 12 cm long, with numerous small
leaflets and distinctly but narrowly winged rachis. A young tree of
this species was observed growing very vigorously in Colonel Robert H.
Montgomery's arboretum at Coconut Grove, Florida, in the peculiar sandy-loam
and porous-limestone soil characteristic of the extreme southeastern section of
that state. The long, nearly straight side branches were extremely
spiny. Because of its spiny character, this species should be
tested as a hedge plant. As it is clearly related to
Feronia, a genus that forces premature blooming of Citrus grafted
on it, Feroniella should also be tested as a stock for Citrus.
2. Feroniella pubescens Tan.
Bul. Mus. Hist. Nat. Paris, 2 sér. 2:161. 1930.
Type.—Java (Zollinger, No.
493). Herb. Kew (Tanaka ident. No. Q-1197).
Paratype: Burma (Wallich, No. 6365). Herb. Kew.
Distribution (fide Tanaka).—Java,
Burma, Thailand, South Vietnam.
Common
name.—Velvety feroniella.
Tanaka's
original description reads as follows (in translation): "Feroniella pubescens
Tanaka, n.sp. Limonia ? pubescens, Wall. Cat. no. 6365 nom
nud. Twigs spiny, spines small, solitary or paired;
recurved. Leaves odd-pinnate, 5-7-paired, rachis pubescent, more or
less widely winged, leaflets opposite, subsessile, slightly serrulate,
lateral leaflets oblong, acute or subacute, terminal one ovate, acute or
acuminate. Inflorescence axillary, paniculate,
few-flowered. Calyx 5-lobed, lobes somewhat triangular,
pubescent. Petals 5, oblong, acute, puberulous. Stamens
numerous, much longer than the petals, filaments subulate, with a pilose
appendage, anthers somewhat rounded. Style linear, puberulous,
stigma capitate, 5-lobed. Immature fruits small, obovate, with 5
locules, cortex woody, of homogenous structure."
Tanaka made a specimen collected in Java by
Zollinger (No. 493, in the Kew Herbarium) the type of this species, but he
considered Wallich's No. 6365, collected in Burma at Taong-dong in 1826 and
preserved in the Kew Herbarium, as a paratype. He provided Swingle
with a photograph of the paratype. Wallich's specimen was studied
by J. D. Hooker (1875, p. 507), who found only three to five pairs of leaflets
(not five to seven, as Tanaka stated). The lateral leaflets are 3.5
to 6 by 0.8 to 12 cm, with the margins subentire or slightly and irregularly
serrulate above the middle but entire and cuneate below the middle; the rachis
segments are 1.2 to 2.5 cm long, with rather broad wings, 2 to 4 mm
wide. This specimen looks like a form of Hesperethusa
crenulata with the leaflets less crenullate than usual. The
fruit that Wallich associated with this number was suspected by Hooker (1875,
vol. 1, p. 508) of being "probably that of another plant." Tanaka
described an immature, 5-locular, obovate fruit with a woody shell, of
homogenous texture that does not fit well with Feroniella as now
known. However, he described flowers that have many stamens with
pilose appendages on the filaments, such as are known only in
Feroniella.
Until better material can
be studied, the true character of this species must remain in doubt.
3. Feroniella lucida (Scheff.)
Swing. Bul. Bot. Soc. France 59:781. 1912. Feronia lucida Scheff.
1870. Illus., Koorders & Valeton, Icon. Bogor. 2:190, pl. 149.
1904; fig. 3-58 this work.
Type.—Java, Rembang Province (Teijsmann). Herb. Hort. Bot.,
Buitenzorg (now Kebun Raya, Bogor, Java).
Distribution.—Central Java; in teak (Tectona grandis) forests.
Common name.—Java feroniella.
The fullest description yet made of this species
is that of Koorders and Valeton (1896, pp. 251-53); it reads in translation as
follows: "Twigs slender, gray, young tips pubescent. Leaves 2-5-
(usually 3-4-) paired; rachis pubescent, articulate at the nodes, internodes
more or less winged above, petioles very short, pubescent. Leaflets
elliptic or obovate, obtuse at the apex, often cuneate at base, entire or
subcrenulate, margins recurved, shining and glabrous above, below opaque
pubescent all over or only at the veins, pellucid-punctate over the whole
surface, but having a single series of larger glands near the
margin. Panicles loose, made up of 3-flowered cymes, often 2 or 3
grouped in the axils, pubescent, shorter than the leaves. Flowers
large, hermaphrodite, or staminate by abortion of the stigma and ovules (the
ovary seems well developed and grows for a time); flower-buds opening early
with all [flower] parts continuing to grow. Calyx lobes linear,
pubescent, deciduous; petals acuminate, margins reflexed at base, opening
erect, then recurving, glabrous; stamens 16-25 in mature flowers, exserted;
anthers oblong, 4-cornered, cordate at the base, truncate-mucronulate at the
apex; pollen [grain] round with 3-4 pores, with granulose exine; ovary globose,
glabrous; style in perfect flowers elongate, terete, glabrous with an exerted
persistent fusiform stigma; in the majority of the flowers (male?) the style is
very short, the stigma deformed or aborted. Fruits with a very
thick woody epicarp. Seeds immersed in glutinous pulp arising from
the placenta and the endocarp; testa thin, woody, with a coriaceous
tegment.
"Small or medium-sized
trees. Leaves 60-100 mm long, internodes 10-20 mm; leaflets 22-30 mm
long, 15-22 mm wide; petiolules 1-2 mm. Petals in fully developed
flowers 12-14 mm; stamens 10-13 mm; disk villose, 2-5 mm high; anthers 3
mm. Ovary with style 13 mm. Fruit 60-70 mm diam.;
epicarp 10 mm thick."
This species is native
in central Java, usually growing at less than 400 meters altitude.
It is a medium-sized tree, 10 to 15 meters (30 to 49 feet) tall, with a
straight trunk 20 to 30 cm in diameter. It is common in stands of
teak (Tectona grandis). It sometimes occurs in situations
where the soil is periodically very dry, for example, at Kedoengdjati in the
Semarang area.
Koorders and Valeton (1896, pp.
251-53) first discovered that in this species, as in Feronia limonia, a
large part of the flowers are male through partial abortion of the
ovary. Feroniella lucida is called kawista krikil by
the Javanese, who eat the fruit as they do that of the cultivated Indian wood
apple, Feronia limonia, which is called kawista by them.
The young trees of this species (see fig. 3-58), like those of the Indo-Chinese Feroniella
oblata, have slender, straight branches armed with numerous long, slender,
sharp thorns frequently arising in pairs. It should be tested as a
hedge plant and as a stock for dwarfing Citrus. Experiments
made by the U.S. Department of Agriculture in greenhouses in the Washington,
D.C. area have shown that Citrus, although it grows very slowly on this
rootstock, may be forced into early blooming by this stock, as it is by the
related Indian wood apple, Feronia limonia. Feroniella lucida
should, like F. oblata, grow well in extreme southeastern Florida.
RETROSPECT AND PROSPECT
In concluding this discussion of the taxonomy of
the orange subfamily, it may not be amiss to emphasize the practical as well as
the scientific importance of the wild relatives of our cultivated citrus
trees. Probably no other important cultivated crop plants can boast
such a significant and varied group of wild relatives. The plants
allied to Citrus show all degrees of affinity with our cultivated
oranges, lemons, and grapefruits. Some of them are to be found in
the genus Citrus; others are in genera closely related to it; still
others belong to distantly related tribes, which may show little superficial
resemblance to Citrus.
The
Citrus wild relatives exhibit a wide range of adaptability to climatic
and soil conditions. Mention of but a few of the more outstanding
examples should suffice to substantiate this statement.
Eremocitrus, of semiarid habitat, has the ability to grow with little or
no organic nitrogen in the soil; it also withstands relatively high
concentrations of salts in the soil moisture. Severinia
endures percentages of boron in the soil solution strong enough to kill
Citrus; even more striking is the fact that under such conditions
Severinia roots absorb and transmit so little boron that they have
supported experimentally healthy grafts of even the highly sensitive
lemon. Other wild relatives of Citrus have acquired,
through long ages of natural selection, high resistance to the attacks of
certain insect pests or fungous diseases. A species of
Citropsis, native to Republic of the Congo, is immune not only to the
attacks of a beetle whose larvae burrow in the collar of Citrus trees,
but also to the foot rot fungus that is made more destructive to Citrus
by the injuries caused by the beetle. This Citropsis (C.
gilletiana) has been found to make a good rootstock in Republic of the Congo
for several cultivated species of Citrus, which grow and fruit well when
grafted on it; however, it cannot be grown on any Citrus rootstock in
that country.
The promising beginning that has
been made in using these wild relatives, and also hybrids derived from them,
as rootstocks for our cultivated varieties of Citrus opens up
possibilities in citrus culture well worth the attention of citrus
experts. To study and test to the best advantage these wild
relatives of Citrus, it is essential to have a thorough knowledge of
their taxonomic relationships as well as of their physiological peculiarities
and special adaptations to the climatic and soil conditions of their native
habitat. In addition to the species of wild relatives and their
hybrids that are useful as rootstocks, there are others closely enough related
to the species of Citrus to hybridize with them and yield promising new
types.
The hybridization of the wild species
of True Citrus Fruit Trees with the cultivated forms has also opened a new
field of research on the origin of some of the more puzzling cultivated
varieties. The Yuzu, a hardy variety widely grown in Japan and
sparingly in northern China as a substitute for the lemon, has been considered
by some to be a good natural species of Citrus. However, it
is now found to be a hybrid of a recently discovered Chinese species of
Citrus (C. ichangensis) probably with a sour mandarin (C.
reticulata var. austera). A hybrid of similar parentage
made in the United States is astonishingly like the Yuzu.
The study of Citrus and its wild
relatives has already yielded appreciable results and it is gratifying to know
that botanical exploration is very likely to bring to light additional new
species or even new genera of Citrus relatives. The most
recent general treatise describing all the genera and species in the orange
subfamily except Citrus was published in 1861 by Daniel Oliver, who
recognized twelve genera and fifty species, some of them doubtful.
As late as 1896, Adolph Engler recognized and described only fourteen genera
and estimated the number of species at seventy-one. In contrast to
this slow increase in the number of genera and species recognized during the
second half of the nineteenth century, a very rapid increase has occurred since
1900. The present synopsis recognizes no fewer than thirty-three
genera and 203 species, of which seventeen genera and eighty species have been
described since 1900, most of them since 1910. In addition to these
eighty new species, some fifty species named before 1900 but not well
understood have been studied anew and are now accepted as valid. In
other words, seventy-five years ago nearly two-thirds of the species recognized
today were either completely unknown or not known adequately.
Vast areas of the Monsoon region, including the
most densely populated part of the distribution area of the orange subfamily,
are still inadequately known. When these regions and certain
sections of tropical Africa are adequately explored by competent botanical
collectors we may expect new species and even some new genera of the orange
subfamily to be brought to light.
Doubtless
many physiological peculiarities, as yet undiscovered, are possessed by certain
of the wild relatives of Citrus, peculiarities that have arisen during
the several tens of million of years in which these plants have been evolving
independently and becoming more and more fully adapted to the special
environmental conditions prevailing in the areas inhabited by the respective
species. These areas extend from the near-desert of east-central
Australia through the tropical jungles of the Monsoon region to the semiarid
foothills of Northwest India and West Pakistan; reappearing again in Africa,
they extend from the mountains and plateaus of East Africa to the dense jungles
of the lower Congo and the lands bordering the Gulf of Guinea in West
Africa.
In the face of such abundant material,
citrus experts can undoubtedly solve certain important technical problems with
greater ease if the varied behavior of the numerous near and distant
Citrus relatives is seen in proper perspective.
Looking ahead, it is reasonable to expect that
citrus breeders will use wild relatives of Citrus, not only as parents
for new types of disease-resistant or otherwise superior citrus fruits and for
breeding new hybrid rootstocks adapted to special soil conditions or resistant
to root diseases, but also as research material to aid in the solution of many
difficult questions in the phylogeny of our cultivated species of
Citrus. Through experimentation and research, taxonomists,
floral anatomists, cyto-taxonomists, and chemo-taxonomists will develop better
explanations concerning the nature and relationship of citrus taxa.
The taxonomic system will be revised and improved. The truth will
be approached but it will never quite be attained because man's knowledge
concerning living organisms forever remains incomplete.
ACKNOWLEDGMENTS
The acknowledgments
presented below were made by the senior author, Walter T. Swingle, in the first
edition of The Citrus Industry (Webber and Batchelor, 1943, vol. 1, pp.
128-29). The acknowledgments are published here without change as
they originally appeared.
1. To
Mr. Benjamin Y. Morrison, in charge of the Division of Plant Exploration and
Introduction, for putting at my disposal all the resources of his division,
both of material and apparatus and of personnel.
2. To the following persons and
institutions for the loan of herbarium specimens, photographs of type
specimens, notes, etc.: Sir Arthur W. Hill, Director of the Royal Botanic
Gardens, and Mr. H. C. Sampson, Economic Botanist, Kew, Surrey, England; Dr,
Edmund G. Baker, Department of Botany, Museum of Natural History, South
Kensington, London, England; Mr. R. E. Holttum, Director of the Botanic
Gardens, Singapore, Straits Settlements; Mr. I. Henry Burkill, Former Director
of the Botanic Gardens, Singapore, Straits Settlements, and now working at the
Royal Botanic Gardens, Kew, Surrey, England; Professor J. R. Matthews, Regius
Professor of Botany and Curator of the Garden, University of Aberdeen,
Aberdeen, Scotland; Dr. H. J. Lam, Director of the Rijks Herbarium, and Dr. S.
J. van Oosterstroom, First Assistant, Leiden, Holland; Professor W. Robyns,
Director of the National Botanic Garden, Brussels, Belgium; Professor N. E.
Svedelius, Director of the Botanical Institute, and Dr. Carl G. Alm, keeper of
the Hortus Linnaeanus, University of Uppsala, Sweden; Professor Dr. L. Diels,
Director of the Botanical Gardens and Botanical Museum, and Dr. M. Domke,
Associate Curator of the Herbarium, Berlin-Dahlem, Germany; Professor Henri
Humbert, Director of the Laboratory of Botany (Phanerogams), Museum of Natural
History, Paris, France; Professor Cyril T. White, Government Botanist of
Queensland, and Director of the Botanic Garden, Brisbane, Australia; Professor
Tyôzaburô Tanaka, Director of the Horticultural Institute, Taihoku Imperial
University, Taihoku, Formosa, Japan; Dr. Eduardo Quisumbing, Curator of the
Herbarium, Bureau of Science, Manila, P.I.; Mr. K. Biswas, Director of the
Royal Botanic Garden, Calcutta, India; Dr. Elmer D. Merrill, Arnold Professor of
Botany, Administrator of the Botanical Collections of Harvard University, and
Director of the Arnold Arboretum, Jamaica Plain, Massachusetts; Dr. Herbert L.
Mason, Associate Curator of the Herbarium, University of California, Berkeley,
California; Dr. H. A. Gleason, Curator of the Herbarium, and Dr. A. C. Smith,
Associate Curator, New York Botanical Garden, Bronx Park, New York City; Dr.
William R. Maxon, Curator of the U. S. National Herbarium, U. S. National
Museum, Washington, D.C.
3. To Dr.
Albert H. Tillson, Junior Botanist, Bureau of Plant Industry, Plant
Introduction Garden, Glenn Dale, Maryland, for efficient cooperation in using
and improving the Juel method of restoring herbarium specimens of flowers,
fruits, etc., and making serial microtome specimens of them.
4. To my wife, Maude Kellerman
Swingle, who aided me for several years in the preparation of this chapter and
who is joint author with me of all the taxonomic names used in the genus
Citropsis.
FOOTNOTES
1. The Monsoon region is used here in a broad sense to include southeastern Asia, the East Indian Archipelago, New Guinea, Melanasia, northeastern Australia, and New Caledonia.
2. The specific name excavata was no doubt employed by Burmann because of the hollowed out shape of the base of the filaments.
3. C. Lauterbach (1918, p. 221) gives the Aurantioideae as having an altitudinal limit of about 300 meters in New Guinea, but reports that several genera of the Toddalioideae and Xanthoxyleae are found growing in cloud and fog belts of the mountains at 800 to 1,500 meters altitude and that three species of Acronychia occur at altitudes of from 2,700 to 3,300 meters.
4. These protuberances when restored nearly to their original dimensions by soaking in hot water and dilute ammonia water are 0.9 to 1.2 mm in diameter. Serial microtome sections show a central woody cylinder 0.5 to 0.6 mm in diameter, with pith in the center and numerous (about 120 to 130) radial rows of thick-walled wood parenchyma cells (4 to 8 rows deep) that are separated from the larger thinner-walled cells of the peripheral tissues by very thin-walled cambium-like cells. On the inner boundary of the woody cylinder next to the pith are found eight groups (each of 4 to 10 cells) of wood vessels with still thicker, pitted walls. The epidermal cells are unusually large; no oil glands were seen. Longitudinal sections showed cells longer (some very much larger) than broad and almost all with cross walls running at right angles.
5. The Latin name of this species was, by some accident, omitted from the original description published in Nos. 4 and 5, October 1, 1911, and December 20, 1911, and did not appear until more than two years later in No. 12 (in the index on p. 390 and in the "Errata et Corrigenda" on p. 406), published on December 31, 1913.
6. The first publication by Wallich of Buchanan-Hamilton's generic name was in the form Lavanga, as nearly as can be deciphered from the minute lithographed manuscript writing. This was apparently a mistake, as the vernacular name is derived, according to Roxburgh (1832, vol. 2, p. 380), from the Sanskrit name luvunga-luta (called a Bengali name by Voigt, 1845, p. 140).
7. H. H. Hume was the first citrus expert to utilize pulp-vesicles as an important means of distinguishing the commonly cultivated species of Citrus Fruit Trees. He states (1909, p. 14; also 1926): "It is believed that the juice sacks of the pulp, their size, shape and appearance in the cut surface of the fruit, can be used to some extent as a diagnostic feature in classification, a point which seems to have been quite generally overlooked." Hume figured the pulp-vesicles and used them in his technical descriptions of the eight species of Citrus, Fortunella, and Poncirus that he recognized.
8. Roemer was apparently the first to notice that the Limonia monophylla of Loureiro (1790, p. 271), wrongly credited by Loureiro to Linnaeus, was not that species but the very different plant, Severinia buxifolia, which Roemer renamed in Loureiro's honor.
9. Tanaka repeated Loureiro's error and wrongly identified Limonia monophylla L. with Severinia buxifolia because he found poor specimens of this latter species labeled Limonia monophylla in the herbarium of Linnaeus, preserved by the Linnean Society of London. However, B. Dayton Jackson, for many years the keeper of the Linnean Herbarium, had already indicated categorically (1912, pp. 26 and 27) that of the material in the Linnean Herbarium labeled Limonia monophylla (including the specimens seen by Tanaka) "there is no specimen so named by Linné," a verdict confirmed by Airy-Shaw (1939, p. 292).
10. This name is given by Groff (1923, p. 273) in his list of some 7,600 Chinese plant names used in Kwangtung Province, China. The Chinese characters used to write this name are given by Eitel and Genähr (1911, characters Nos. 6871, 5047, and 3286), with the tones accurately marked.
11. A curious plant was described under the name Lamiofrutex by C. Lauterbach (Nova Guinea 14:147 [1924]) as a Rutaceae of doubtful position because no fruits were known. This plant is very distinct from any other genus of the Aurantioideae because (1) the ovules (single in the unilocular ovary) are erect; (2) the leaves have very few lateral veins, have wavy margins, are decurrent at the base, and are somewhat shining below; (3) the stems are very slender, creeping and rooting, sparingly branched; and (4) its habitat is in the fog forest at 1,420 to 1,450 m altitude.
The authors have omitted it from the Aurantioideae, although it was included by Engler (1931, p. 330) immediately after Atalantia (used by him in a broad sense) in his last treatise on the Rutaceae. A photograph of the type specimen, kindly sent Swingle by the Director of the Botanical Museum at Berlin-Dahlem, shows leaves very different from those of any species known in the orange subfamily.
12. Made from herbarium materials by the modified Juel method.
13. Ripe fruits of H. crenulata that Swingle picked on May 30, 1939, from a tree growing at the Horticultural Experiment Station at Deodoro near Rio de Janeiro (a plant which Swingle had sent to Brazil through Snr. Felisberto de Camargo about ten years before), were very bitter, not sour. Probably the oft-recurring statement that the fruits of this species are "very acid" stems from the general but improper attribution to it of the name Limonia acidissime L., a name originally applied by Linnaeus to a very different plant, the wood apple, here called Feronia limonia (see. p. 416), the fruit of which is said to have "a strong acid-sweet taste" and to make excellent jelly.
14. Some of the species of Atalantia, such as A. ceylanica, have fruits with very few pulp-vesicles, apparently because the seeds have become very large and fill the locules of the fruit almost completely, leaving little room for pulp-vesicles. These species constitute the subgenus Rissoa. Another species, A. guillaumini, which has much resemblance to A. ceylanica, seems to have no pulp-vesicles; it has perhaps lost them by retrograde evolution.
15. Usually credited to Wight and Arnott but first published by Wight in January, 1834, with full description and plate, whereas the Prodromus in which it appears as A. racemosa Wt. et Arn. was not published until after March 20, 1834, and furthermore gives only a very brief description scarcely adequate for valid publication.
16. Harry H. Hazelwood, a prominent nurseryman of Epping, New South Wales, reported to Swingle in a letter dated January 23, 1924, that at Carwell Station (Long. 14°7 48' E., Lat. 31° 15' S.), 44 miles southwest of Coonamble, there had been a drought of some months' duration during which a number of trees of the Buddah, Eremophila mitchelli, had died, but that in the large tract covered with a growth of Eremocitrus glauca "not a single instance was noted of these trees dying of drought." O. E. Friend, a resident of Carwell, wrote on November 16, 1915, that the rainfall for the preceding 12 months had been only 9.09 inches and for the year before only 14.80 inches, yet "the trees [of Eremocitrus glauca] were wonderfully green at present in spite of the prolonged droughts." The rainfall at Carwell averages about 19 inches, according to Mr. Friend.
The drought resistance of Eremocitrus glauca was discussed by Flanders (1932), who reported seeing small trees growing at Mootwingee (Long. 142° E.), where the annual rainfall averages about 12 inches. He also told Swingle of seeing living but leafless trees near Longreach, Queensland (Long. 144° E.), where no rain had fallen during the preceding 15 months and no green vegetation could be seen!
17. Tanaka, in his original description and in his later notes (1931b, p. 12) on this plant, reported this species as native to New Guinea. He was evidently in error, as the type specimen only (Peekel, No. 408) is cited, and it was collected in New Ireland, some 600 kilometers northeast of the nearest shore of New Guinea.
18. P. Brough (1933, p. 68) cited W. N. Benson (1923, pp. 1-62) in support of his view that "Australia is considered to have been joined to the Asiatic mainland at least during the Cretaceous period, but probably a complete separation has existed since the beginning of the Eocene." The migrations of higher plants into Australia are held to have occurred during later Cretaceous times.
19. Before its hybrid nature had been established, this form was called, both in the Agricultural Research Service greenhouses at Washington and in the field tests, Microcitrus virgata, a provisional name alluding to its very numerous slender, straight, whiplike twigs. This provisional name was never validated by proper publication, but is sometimes seen in print as, for example, in Fawcett (1936, p. 177).
20. The fifteen "primary elements" of the genera Citrus, Poncirus, and Fortunella given by Tanaka (1935b) are as follows: (1) Citrus macroptera Mont., (2) C. hystrix DC., (3) C. aurantifolia Swingle, (4) C. medica Linn., (5) C. limon Burm. f., (6) C. limonia Osbeck, (7) C. grandis Osbeck, (8) C. aurantium Linn., (9) C. sinensis Osbeck, (10) C. junos Sieb. ex Tanaka, (11) C. ichangensis Swingle, (12) C. tachibana Tanaka, (13) C. depressa Hayata, (14) Poncirus trifoliata Raf., (15) Fortunella japonica Swingle.
21. Tanaka's translation.
22. All species are in fact compatible.
23. There are a number of so-called species of citrus fruit trees that are of doubtful status. They may be natural species of comparatively recent origin, not yet far removed from their ancestral forms, or they may be chance hybrids or sports that have been propagated by man or may even be propagated spontaneously by apogamic embryos that reproduce the hybrid or sport exactly. The term "satellite species" is here used to designate species of doubtful validity that may prove to be one of such forms.
24. As noted, naringin is found in the pummelo (C. grandis) and in the grapefruit (C. paradisi), a satellite species; see also table 3-4.
25. M. B. Matlack (1931, p. 438), in studying the plastids or chromoplasts in the pulp-vesicles of citrus fruits, found that "their size and shape appear to be characteristic of the species from which they are obtained…the tangerine, Satsuma [his fig. 4] and willow-leaved mandarin oranges have spindle-shaped plastids…in the King orange they are isodiametric [his fig. 3]." Such isodiametric chromoplasts occur, according to Matlack, in "the numerous varieties of the sweet orange." In other words, the King orange representing very nearly the type of C. nobilis (as originally published by Loureiro), has chromoplasts similar to those of the the [sic] sweet orange, C. sinensis, and very different from those of the loose-skinned orange, C. reticulata. This is additional proof that it is a hybrid of the sweet orange and the loose-skinned orange.
26. Hall (1938) found in the leaves and in a thin layer of the peel (just between the yellow outer peel and the albedo) a new enzyme of the glycosidase type, able to hydrolize naringin in vitro at low temperatures (37° C.) at pH 7 into "an insoluble aglycone, naranginin (or naringetol)" and a new disaccharide sugar, distinct from rutinose.
27. Unfortunately, the original description covered fruits of the so-called Ichang lemon collected by E. H. Wilson in Hupeh Province. At that time they were believed to be from a cultivated form of C. ichangensis. Seeds of these fruits were introduced into the United States about 1914; trees grown from them have leaves entirely different from the true Ichang papeda. The so-called Ichang lemon is probably a hybrid of C. ichangensis with a species of subgenus Citrus.
28. The moli kana, or edible moli, of the natives, correctly classed by Seemann (1862, p. 321) as C. decumana (= C. grandis).
29. Merrillia, which bears a large fruit with a tough, leathery shell somewhat analogous to that of Swinglea and with seeds covered with flattened hairs (paleae) somewhat resembling the seeds of Swinglea, would seem, at first sight, to belong to this group, but in its habit of growth and in its leaves (with alternate leaflets) and wood it shows close similarity to Murraya paniculata. Swingle followed Engler (1931, p. 211), a lifelong student of the Rutaceae, in removing Merrillia from the subtribe Balsamocitrinae, to which he at first assigned it, and placing it in the tribe Clauseneae next to Murraya.
There are also certain similarities to be seen between Swinglea and Burkillanthus, but the fruit structure of the latter is clearly very different.
30. This unusual number of stamens is almost certainly due to the outer whorl (alternative with the petals) persisting in the ancestral number, whereas the inner whorl is doubled at an early stage of development. Penzig, who held to the view of obdiplostemony in Citrus was hard pressed to find clear proof of any differentiation of the stamens of Citrus into two distinct whorls. He finally discovered (1887, p. 69) a not uncommon monstrosity, a partly double-flowered variety of Citrus in which the stamens of the outer whorl but not the inner whorl were transformed into petals. The flower of A. gabonensis is the first instance yet found in the orange subfamily of a normal (not teratological) difference in the stamens of the two whorls.
31. Airy-Shaw (1939, p. 293) has proposed substituting the generic name Limonia for Feronia. However, in view of the origin and subsequent misuse of the name Limonia such a substitution seems inadvisable. When Linnaeus established the genus Limonia in 1762 he failed to describe it, and although it apparently covered what he assumed to be a single species, Limonia acidissima, he included in the synonomy plants belonging to at least two species, one each, in the genera Hesperethusa and Feronia (citing illustrations of both) and probably to a third species in the genus Citrus! Thus, from the very start, the name Limonia was a nomen confusum, "derived from two or more entirely discordant elements," which "must be rejected" in accordance with Article 70 of the International Code of Botanical Nomenclature (Lanjouw, 1961, p. 50). Furthermore, the name Limonia acidissima was apparently never used by any botanist for the wood apple, but was often misapplied to the plant now known as Hesperethusa crenulata. In 1800 Corrêa da Serra established the genus Feronia for the wood apple, and since that date all botanists have so used it. On the other hand, Limonia has become a true nomen ambiguum, "permanent source of confusion or error," as it has been applied to plants belonging to at least thirteen very diverse genera, namely, Micromelum, Glycosmis, Murraya, Triphasia, Pamburus, Luvunga, Paramignya, Severinia, Pleiospermium, Hesperethusa, Citropsis, Atalantia, and Swinglea, representing both tribes and five of the six subtribes which constitute the subfamily Aurantioideae! In view of the dubious origin and wholesale misuse of the name Limonia the genus name Feronia should be conserved and Limonia should be rejected. For this reason we are using the name Feronia.
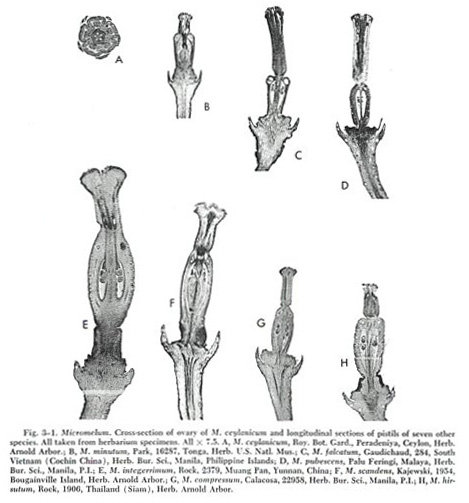{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
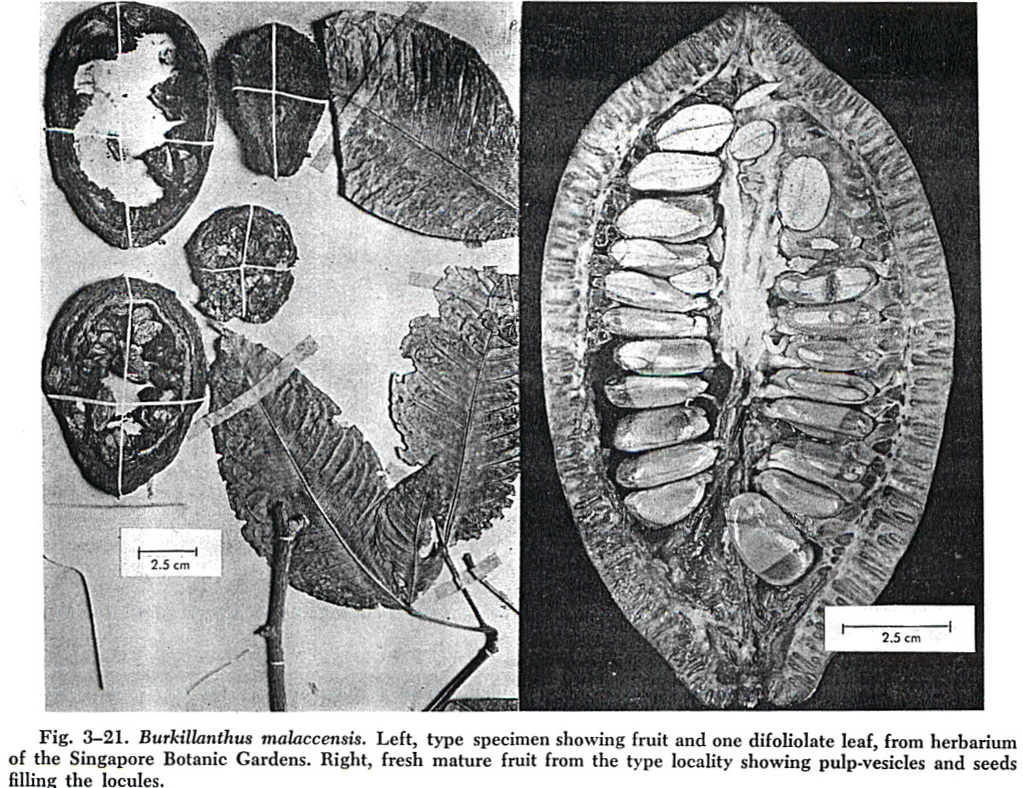{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
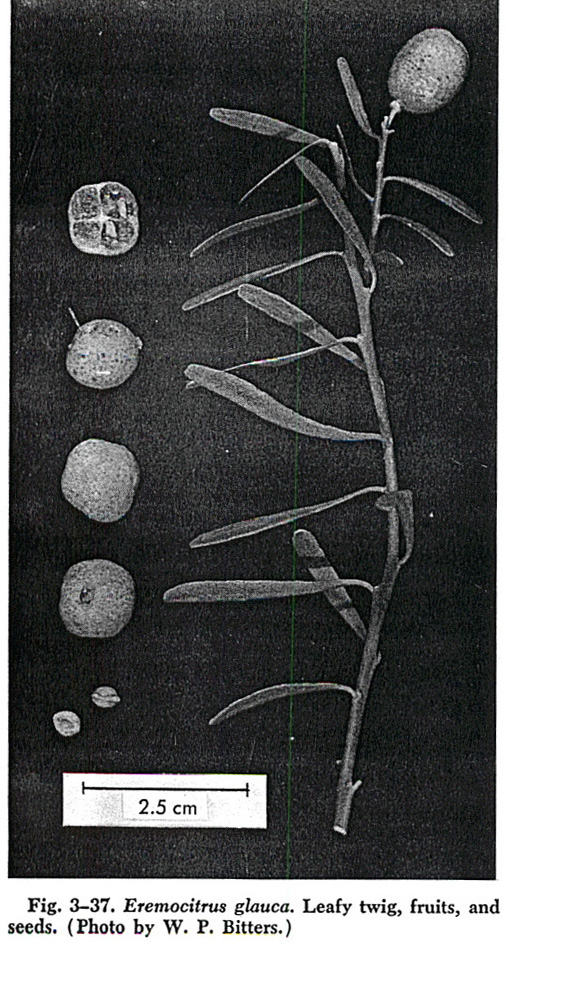{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
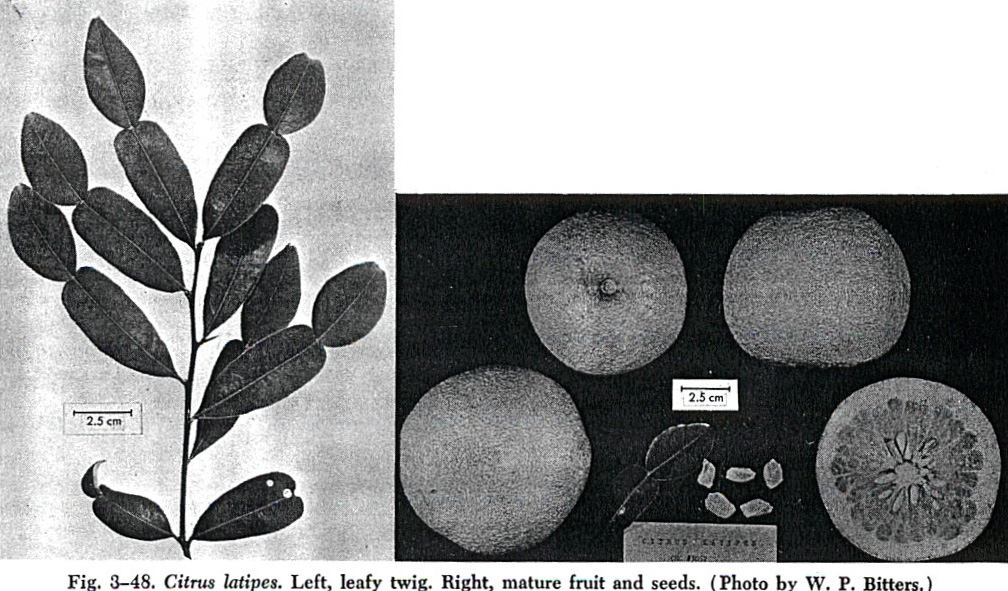{kind=link}
{kind=link}
{kind=link}
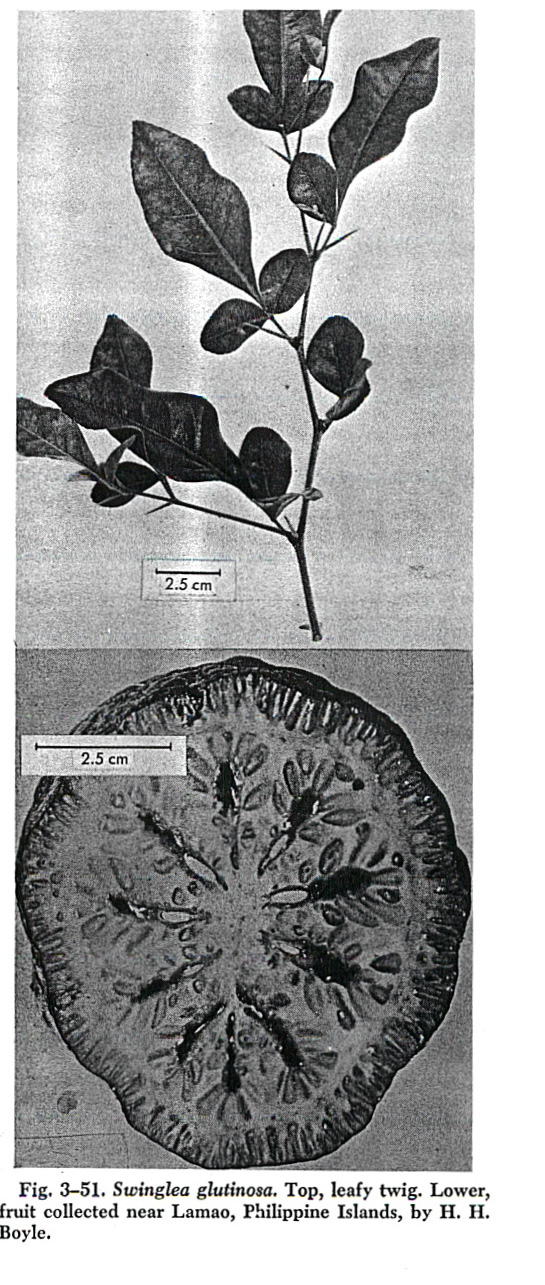{kind=link}
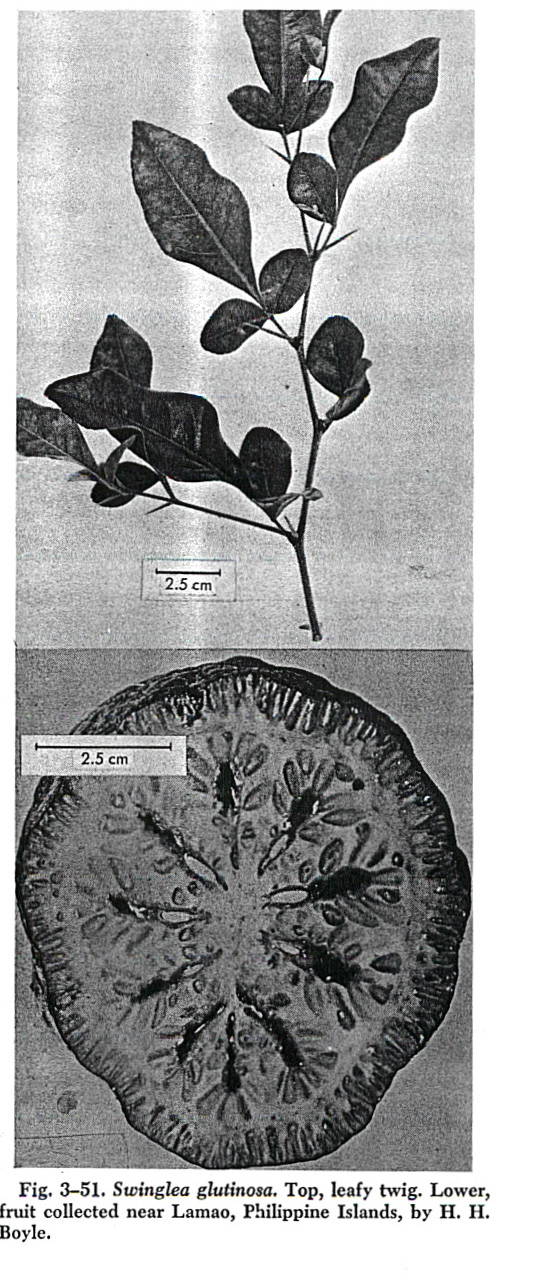{kind=link}
{kind=link}
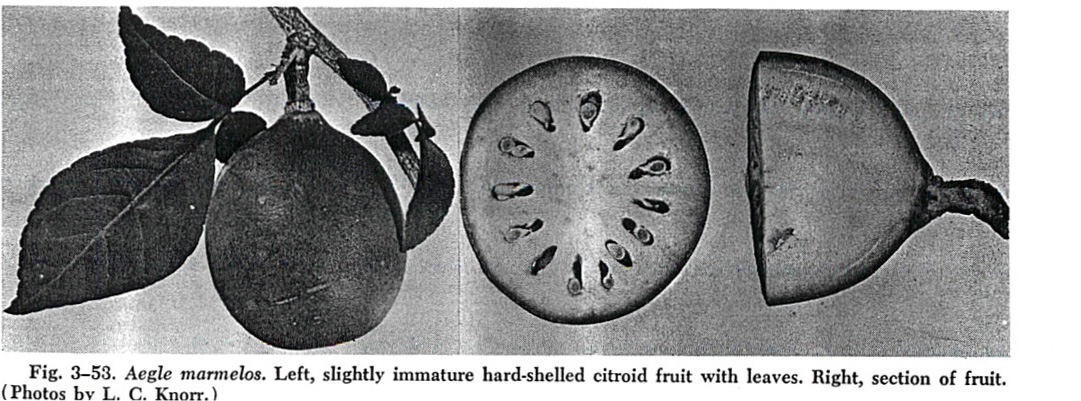{kind=link}
{kind=link}
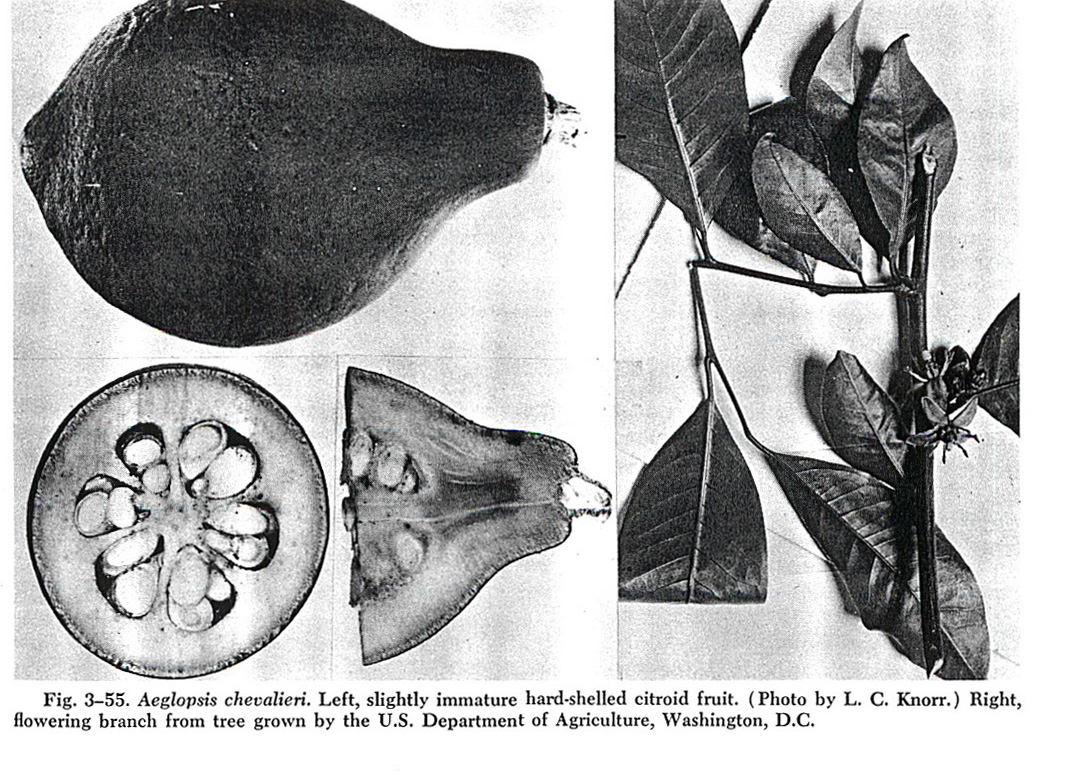{kind=link}
{kind=link}
{kind=link}
{kind=link}